La variación natural en la raíz de la planta más empleada en ciencia: Arabidopsis thaliana
La variación natural es muy importante y para estudiarla ha sido necesario clasificarla en diferentes niveles hasta llegar a la categoría de “especie”, cuya definición biológica es: “aquellos organismos que pueden reproducirse y dar descendencia fértil”. Así, podemos decir que un ahuehuete es una especie diferente que un olmo o que un jaguar es una especie distinta que un león. También hay variación o diferencias entre organismos que pertenecen a la misma especie, lo que se denomina variación natural, que es resultado de la suma de diferencias genéticas y de los distintos ambientes donde habitan diversas poblaciones de una misma especie. Específicamente, se ha estudiado la variación natural de las plantas en diferentes características observables (denominado fenotipo), como el tamaño de las hojas, el tamaño de las raíces o variaciones en el tiempo de floración, por dar algunos ejemplos (Figura 1).

¿Por qué usar a la raíz de Arabidopsis thaliana como modelo de estudio?
Arabidopsis thaliana es la especie de planta más utilizada en la ciencia y pertenece a la misma familia en la que se clasifican la coliflor o el brócoli (Brassicaceae) (Figura 2A). Arabidopsis thaliana se definió como especie en 1842 y un siglo después fue empleada como un organismo modelo para el estudio de la genética y la biología del desarrollo de las plantas (Khanna & Kutschera, 2020). Arabidopsis thaliana es originaria de Europa, Asia y el norte de África (Weigel, 2012), pero se ha introducido a muchos países y actualmente se estudia en todo el mundo.
Arabidopsis thaliana reúne todas las características de un sistema modelo para llevar a cabo proyectos de investigación científica: su tamaño es pequeño, por lo que crece fácilmente en espacios reducidos (Figura 2B); se autopoliniza, lo que permite tener líneas estables; su tiempo de vida es corto (8-12 semanas); una sola planta produce miles de semillas; su genoma es pequeño y se obtienen fácilmente mutantes de esta especie, lo que nos permite estudiar la función de cualquier gen (Khanna & Kutschera, 2020). Debido a lo anterior, es posible realizar estudios en diferentes órganos de esta planta como son las flores, hojas, embriones y raíces.

Las raíces, a pesar de que son la parte oculta de la planta ya que, en general, se encuentran en la tierra, son muy importantes pues le permiten a la planta adherirse al sustrato donde crece; adquieren los nutrientes y el agua del suelo; facilitan la interacción con los microorganismos del suelo y sintetizan varios compuestos orgánicos (Deja-Muylle et al., 2020) (Figura 3A). La arquitectura radicular de Arabidopsis thaliana comprende tanto las raíces laterales como los pelos radicales y la raíz primaria o pivotante, la cual se establece cuando el embrión se está formando.
La raíz primaria está conformada por tres zonas que se localizan en el eje longitudinal: en la parte más apical, es decir, la que tiene contacto con el suelo, se halla el meristemo (el sitio donde las células se dividen o proliferan) de la raíz o RAM (por sus siglas en inglés Root Apical Meristem, Meristemo Apical de la Raíz). Le sigue una zona de alargamiento o EZ (por sus siglas en inglés Elongation Zone, Zona de Elongación) en donde las células se alargan a una tasa muy elevada, y la zona de maduración o MZ (por sus siglas en inglés Maturation Zone) donde las células adquieren sus características finales, como la presencia de pelos (Ivanov & Dubrovsky, 2013) (Figura 3B).
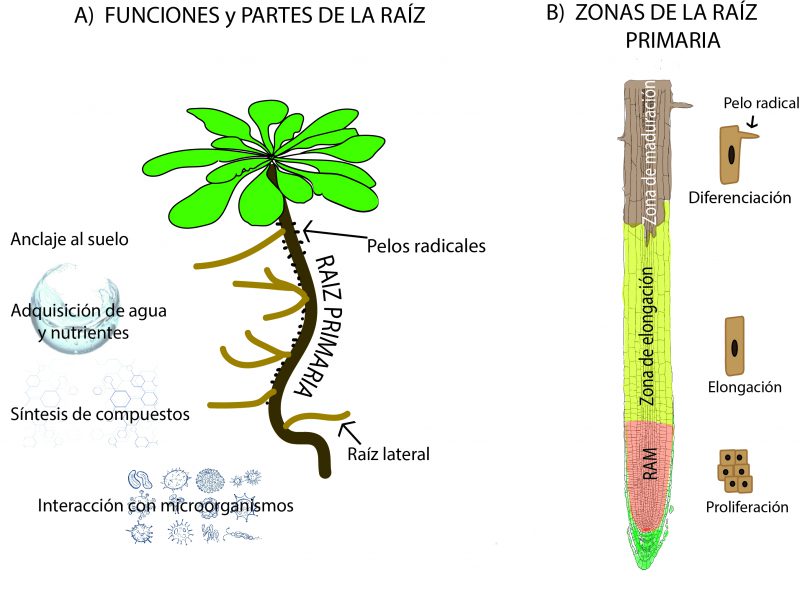
La raíz primaria de Arabidopsis thaliana es un modelo ideal para estudiar la relación entre dos procesos que determinan el crecimiento de cualquier órgano, ya sea animal o vegetal: la proliferación o división de las células y la diferenciación de una célula totipotente, en una célula especializada (Meyerowitz 2002, Dinneny et al., 2008). La proliferación da lugar a nuevas células que alcanzarán su tamaño máximo, y se diferenciarán conforme vayan entrando nuevas células y se desplacen lejos del lugar de la raíz que produce células troncales o madre. Este crecimiento puede estudiarse tanto en condiciones óptimas como en situaciones de estrés, por lo que el estudio de las raíces es relevante para entender la adaptación de las plantas a diferentes ambientes. Además, la raíz es un órgano transparente, con crecimiento continuo, además de tener pocos tipos de células que la conforman (Scheres, & Wolkenfelt, 1998), lo cual facilita su estudio.
Variación natural de Arabidopsis thaliana
Arabidopsis thaliana tiene una amplia distribución en el hemisferio norte con cerca de 7,000 poblaciones naturales que han crecido bajo diversos entornos ambientales. Por ejemplo, crece de forma natural en países tan diferentes como Holanda, Rusia, Tayikistán o Japón y en sitios tan diversos como en un estacionamiento de coches, en la playa, o en lo alto de una montaña. Esto genera diversas variantes naturales de la misma especie a las que se les denomina accesiones, las cuales tienen diferentes nombres; por ejemplo, una de las accesiones que proviene de Japón se llama Sapporo-0 y una de Rusia, Rsch-4. Estas accesiones son una fuente importante de información para identificar genes que participan en el desarrollo de la planta y su respuesta al estrés tanto biótico como abiótico (Alonso-Blanco et al., 2000; Weigel, 2012).
Diferentes condiciones para estudiar la variación natural
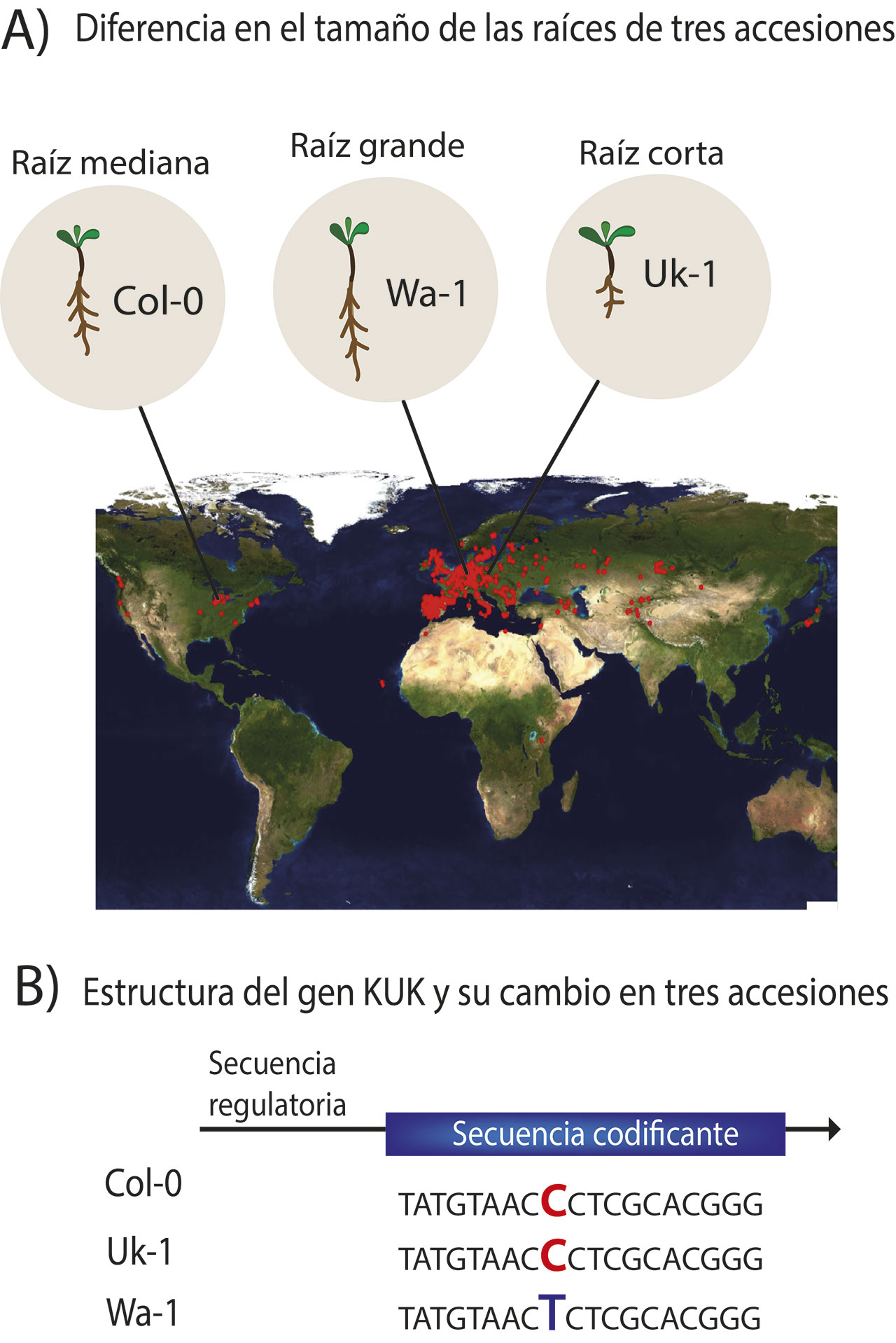
Existen diversas circunstancias en las cuales se puede estudiar la variación natural; por ejemplo, en suelos con diferente salinidad, deficiencia en nutrientes como zinc, nitrógeno, fósforo, hierro, etcétera, o la presencia de metales pesados, entre otros (Deja-Muylle et al., 2020). Además, en condiciones normales de laboratorio -cuando la planta no está sometida a ningún tipo de estrés- es posible ver diferencias muy marcadas en cuanto al crecimiento de la raíz, y pueden clasificarse en aquellas de raíz larga y de raíz corta con respecto a una accesión de referencia, que se usa en casi todos los laboratorios del mundo y que se describió en Estados Unidos como Col-0. De esta manera, se ha visto que la accesión Uk-1 de Alemania que tiene una raíz corta, produce menos células y tiene un meristemo más pequeño, mientras que la accesión Wa-1 de Polonia presenta una raíz larga con más células en el meristemo, que es más grande (Figura 4A) (Meijón et al., 2013).
Por otro lado, se puede estudiar cómo afecta una condición estresante al crecimiento de la raíz; cuando se crece Col-0 en un medio con bajo contenido de zinc, su raíz se desarrolla ligeramente más que en un medio con contenido normal de este mineral. Contrario a esto, la raíz de Sapporo-0, de Japón, crece mucho menos en el medio con bajo zinc (Bouain et al., 2019). Esto quiere decir que las accesiones responden de forma diferente a una deficiencia de zinc; algunas tienen una raíz más grande que otras. Esto es, pueden usarse diferentes accesiones o poblaciones naturales de la misma especie (Arabidopsis thaliana) para estudiar las adaptaciones a las diferentes condiciones ambientales en las cuales crecieron.
¿Cómo se estudia la variación natural?
Una forma de estudiar la variación natural en cualquier especie, incluidos Arabidopsis thaliana y el ser humano, es mediante una herramienta llamada GWAS (del inglés Genome-Wide Association Studies, Estudios de Asociación del Genoma Completo). Lo que hace este procedimiento es asociar una variante en el DNA de las poblaciones y una característica que pueda medirse para encontrar genes novedosos que participen en un fenotipo determinado; en nuestro caso, la longitud de la raíz primaria. Por ejemplo, una secuencia de DNA de un gen puede presentar un cambio en una o más letras, lo cual se asocia a raíces más largas en ciertas poblaciones, pero no en las accesiones con raíces cortas (Figura 4B). De esta forma, el GWAS realiza una asociación entre una secuencia de DNA y una característica que fue medida en cientos de accesiones, comúnmente más de 100 colectadas en diferentes partes del mundo. Con el GWAS se pueden encontrar genes nuevos que intervienen en una característica en particular, como el tamaño de la raíz (Bush & Moore, 2012) o encontrar variantes de genes que pueden ser más propensas a enfermedades como el cáncer.
De las accesiones Uk-1 de Alemania, con raíz y meristemo corto y Wa-1 de Polonia, con raíz y meristemo grande, empleando GWAS se encontró un gen llamado KURZ UND KLEIN o KUK que en alemán significa corto y pequeño. Este gen presenta cambios en la región codificante (es decir la secuencia de DNA que originará la proteína funcional) en las accesiones que tienen meristemo grande, incluida Wa-1, mientras que no hay estos cambios en las accesiones con meristemo corto como Uk-1 (Figura 4B) (Meijón et al., 2013). Esto significa que, cambios en la secuencia codificante de este gen son suficientes para que la raíz de Wa-1 crezca más. También se encontró que KUK se expresa más en el meristemo de la raíz de Wa-1 y menos en el de Uk-1, lo que quiere decir que el producto de este gen es requerido en mayor medida para el crecimiento de la raíz (Meijón et al., 2013). De esta forma y mediante el uso de GWAS, se encontró un gen nuevo que participa en el tamaño de la raíz. Hasta el momento no se ha realizado algún estudio que asocie las variaciones en este gen a diversos tipos de estrés, incluido el estrés por deficiencia de zinc; sin embargo se ha reportado que versiones del gen KUK en donde hay nula producción de la proteína están asociadas a la adaptación de accesiones de Arabidopsis thaliana a la cuenca del Rio Yangtze en China, con un clima muy diferente al de su lugar de origen, que es Europa (Xu et al., 2019).
Conclusiones
Estudiar la variación natural usando la raíz de la planta modelo Arabidopsis thaliana ha permitido entender las enormes diferencias fenotípicas de una población y comenzar a dilucidar algunos mecanismos moleculares que subyacen al crecimiento de las mismas. Estas diferencias fenotípicas reflejan adaptaciones a diversos medios ambientes que, en muchos casos resultan estresantes, como pueden ser la disminución de nutrientes, la presencia de metales pesados o la alteración en las condiciones del suelo como la salinidad, por mencionar algunos. Una de las herramientas que nos ayuda a estudiar esta variación natural es el GWAS ya que posibilita la localización de genes novedosos a partir de la asociación del DNA con una característica fenotípica que, en nuestro caso, es la longitud de la raíz.
Encontrar nuevos genes que alteren los procesos de proliferación/diferenciación celular en respuesta a diferentes estímulos ambientales es clave para entender el crecimiento de cualquier órgano y explicar diversos aspectos de la evolución, el desarrollo y la genética de los organismos vegetales. La raíz de Arabidopsis thaliana resulta un modelo ideal para llevar a cabo este tipo de estudios.
Agradecimientos
Se agradece la ayuda de la Dra. Diana Belén Sánchez Rodríguez por su apoyo en el laboratorio, y al Dr. José Gerardo Rodríguez Tapia de la Unidad de Geomática del Instituto de Ecología, UNAM por la elaboración de los mapas.
Referencias
Alonso-Blanco, C., & Koornneef, M. (2000). Naturally occurring variation in Arabidopsis: an underexploited resource for plant genetics. Trends in plant science, 5(1), 22-29. DOI: 10.1016/s1360-1385(99)01510-1
Bouain, N., Korte, A., Satbhai, S. B., Nam, H. I., Rhee, S. Y., Busch, W., & Rouached, H. (2019). Systems genomics approaches provide new insights into Arabidopsis thaliana root growth regulation under combinatorial mineral nutrient limitation. PLoS Genetics, 15(11), e1008392. DOI: 10.1371/journal.pgen.1008392
Bush, W. S., & Moore, J. H. (2012). Genome-wide association studies. PLoS computational biology, 8(12). DOI: e1002822. 10.1371/journal.pcbi.1002822
Cajero-Sanchez, W., Aceves-Garcia, P., Fernández-Marcos, M., Gutiérrez, C., Rosas, U., García-Ponce, B., … & Garay-Arroyo, A. (2019). Natural Root Cellular Variation in Responses to Osmotic Stress in Arabidopsis thaliana Accessions. Genes, 10(12), 983. DOI: 10.3390/genes10120983
Deja-Muylle, A., Opdenacker, D., Parizot, B., Motte, H., Lobet, G., Storme, V., … & Beeckman, T. (2022). Genetic variability of Arabidopsis thaliana mature root system architecture and genome-wide association study. Frontiers in Plant Science, 12, 3246. DOI: 10.3389/fpls.2021.814110
Dinneny, J. R., & Benfey, P. N. (2008). Plant stem cell niches: standing the test of time. Cell, 132(4), 553-557. DOI: 10.1016/j.cell.2008.02.001
Ivanov, V. B., & Dubrovsky, J. G. (2013). Longitudinal zonation pattern in plant roots: conflicts and solutions. Trends in Plant Science, 18(5), 237-243. DOI: 10.1016/j.tplants.2012.10.002
Khanna, R., & Kutschera, U. (2020). Arabidopsis: two-hundredths anniversary of its name and the possibility of a hidden universal regulatory signal. Journal of Plant Biochemistry and Biotechnology, 29(4), 575-579. DOI: 10.1007/s13562-020-00609-1
Korte, A., & Farlow, A. (2013). The advantages and limitations of trait analysis with GWAS: a review. Plant methods, 9(1), 29. DOI: 10.1186/1746-4811-9-29
Meijón, M., Satbhai, S. B., Tsuchimatsu, T., & Busch, W. (2014). Genome-wide association study using cellular traits identifies a new regulator of root development in Arabidopsis. Nature genetics, 46(1), 77-81. DOI: 10.1038/ng.2824
Meyerowitz EM. 2002. Plants compared to animals: the broadest comparative study of development. Science. 295(5559):1482-5. Review.
Scheres, B., & Wolkenfelt, H. (1998). The Arabidopsis root as a model to study plant development. Plant Physiology and Biochemistry, 36(1-2), 21-32. DOI: 10.1016/S0981-9428(98)80088-0
Soto-Cerda, B. J., & Cloutier, S. (2012). Association mapping in plant genomes. Genetic Diversity in Plants, 29-54. 10.5772/33005
Weigel, D. (2012). Natural variation in Arabidopsis: from molecular genetics to ecological genomics. Plant physiology, 158(1), 2-22. DOI: 10.1104/pp.111.189845
Xu, Y. C., Niu, X. M., Li, X. X., He, W., Chen, J. F., Zou, Y. P., … & Guo, Y. L. (2019). Adaptation and phenotypic diversification in Arabidopsis through loss-of-function mutations in protein-coding genes. The Plant Cell, 31(5), 1012-1025.