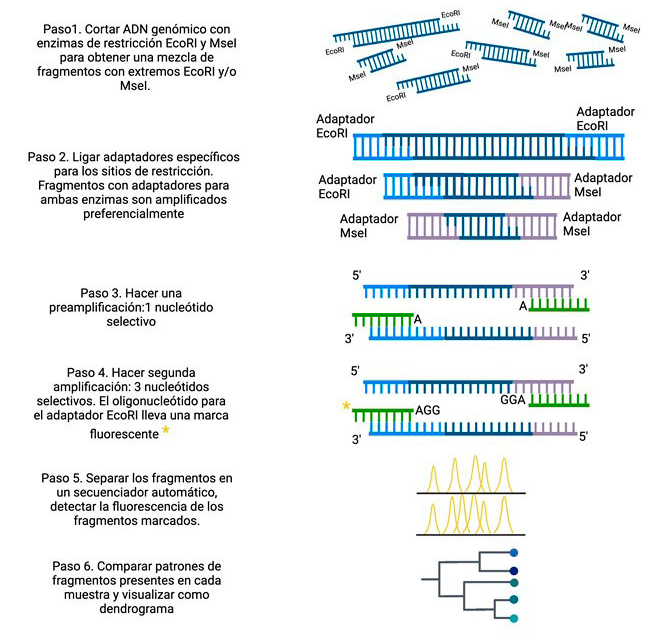
Figura 1. Representación gráfica de la respuesta calorimétrica de vesículas biohíbridas (BhVs) compuestas de fosfolípidos, proteínas y copolímeros; y su comparación con liposomas unilamelares (LUVs) de DMPC y DMPC/LYZ. Las curvas de la gráfica fueron obtenidas por calorimetría (DSC) y representan la respuesta térmica de los sistemas simples (LUVs) y de los sistemas biohíbridos compuestos de DMPC (fosfolípido), LYZ (proteína) y copolímeros de PEGA-b-BuA y PEGA-b-LA. La ilustración representa una sección de una vesícula biohíbrida con sus respectivos componentes.
Motivadas por el desarrollo de terapias farmacológicas más eficientes, la industria farmacéutica, la nanomedicina y áreas relacionadas, se encuentran inmersas en una carrera para encontrar los mejores “caballos de Troya” y así controlar, de manera más efectiva, la administración de fármacos para el cuidado de la salud humana. En la actualidad, la investigación en nanomedicina ha ido en aumento y detonado la era de los sistemas inteligentes para el transporte y liberación controlada de fármacos. En este contexto se han propuesto diversos nanosistemas, con el objetivo principal de lograr terapias farmacológicas más eficaces, disminuyendo los efectos no deseados.
Cabe resaltar que los mecanismos de acción de un fármaco, en un entorno de terapia farmacológica, se basan en el clásico mecanismo propuesto de “llave-cerradura”, lo cual es una forma sencilla de ejemplificar las interacciones específicas. Sin embargo, dada la naturaleza fisicoquímica de cualquier fármaco, éste no solo está sujeto a interacciones específicas en el cuerpo de un organismo vivo, sino que también está expuesto a todas aquellas interacciones de tipo no específicas, es decir, el fármaco puede interaccionar con otros componentes bioquímicos durante su administración que no corresponden al blanco terapéutico de interés, disminuyendo así la eficiencia de la terapia farmacológica. De este modo, no podemos imaginar que un fármaco, por más específico que sea, lleve a cabo su acción de manera eficiente si no lo protegemos de ese constante “bombardeo” de interacciones no específicas.
Inspirados por estos avances y con una extensa colaboración interdisciplinaria, interinstitucional e internacional, investigamos las propiedades termodinámicas y morfológicas de vesículas biohíbridas (BhVs) compuestas de lípidos, proteínas y polímeros propuestas como sistemas potenciales para el acarreo de fármacos. Estos sistemas fueron ensamblados utilizando DMPC como componente lipídico, LYZ como componente proteico y copolímeros de PEGA-b-BuA y PEGA-b-LA como constituyentes poliméricos. Aunado a ello, para evaluar el potencial de estos sistemas también empleamos un fármaco modelo: tetracaína (TC), un anestésico local con propiedades lipofílicas y afinidad a las membranas lipídicas, que se utiliza para inhibir el dolor en algunas intervenciones médicas. Las propiedades fisicoquímicas de las BhVs con y sin TC fueron investigadas por medio de calorimetría diferencial de barrido (DSC) y dispersión dinámica de luz (DLS). Nuestros resultados revelan que LYZ y los copolímeros se adsorben en la membrana lipídica cambiando así sus propiedades termodinámicas. Los sistemas que contienen BuA tienden a desestabilizar la membrana lipídica aumentando el índice de polidispersidad de los liposomas. Además, destaca que nuestros resultados revelaron que la tetracaína no tan solo puede fluidizar a las membranas lipídicas, sino que, en combinación con los copolímeros y lisozima, también puede inducir rigidización en la misma. Nuestra investigación provee información relevante relacionada con las propiedades fisicoquímicas de materiales híbridos que puede ser de mucho interés para el desarrollo de nanosistemas de transporte y liberación controlada de fármacos más eficientes. La trascendencia de esta investigación y nuestros hallazgos, así como los detalles de este trabajo, se pueden consultar en la reciente publicación en la revista BBA – General Subjects [1].
Abreviaturas
DMPC, dimiristoilfosfatidilcolina;
LYZ, lisozima;
PEGA, poli(acrilato de etilenglicol-metileter);
BuA, poli(acrilato de butilo);
LA, poli(acrilato de dodecilo);
Referencia
[1] Pérez-Isidoro, R., Guevara-Pantoja, F. J., Ventura-Hunter, C., Guerrero-Sánchez, C., Ruiz-Suárez, J. C., Schubert, U. S., & Saldívar-Guerra, E. (2023). Fluidized or not fluidized? Biophysical characterization of biohybrid lipid/protein/polymer liposomes and their interaction with tetracaine. Biochimica et Biophysica Acta (BBA)-General Subjects, 1867(2), 130287. https://doi.org/10.1016/j.bbagen.2022.130287.
Pulque: bioproceso ancestral
El pulque es una bebida ancestral de mayor popularidad en México que ha subsistido hasta la actualidad. Esta bebida es viscosa, su contenido de etanol oscila de 4 a 7%, su pH de 3.5 a 4.3, y se produce a través de la fermentación del aguamiel extraída de los agaves (A) pulqueros como la A. salmiana, A. americana, A. mapisaga y de A. hookerii, A. marmorata, A. inaequidens (García-Mendoza, 1998). Los mexicas nombraban a esta bebida según sus propiedades físicas o gustativas como: iztacoctli (pulque blanco), necoctli (pulque mínimamente fermentado), teoctli (bebida de los dioses) y metoctli (vino de maguey). Esta última, al adquirir un olor y sabor desagradable en su proceso de descomposición, era nombrada poliuhquioctli, por lo que fue llamada por los españoles pulque (Cabrera, 1992). Esta bebida en la época prehispánica fue difundida desde la Huasteca hasta Yucatán, y fue de gran importancia para los pueblos indígenas en su dieta diaria y ceremonial (Lappe-Oliveras et al., 2008).
A lo largo de la época colonial, la elaboración y comercialización del pulque fueron fundamentales en la economía mexicana. En 1810 el levantamiento armado de independencia provoca el cese del crecimiento económico. Sin embargo, durante el siglo XIX con el inicio del ferrocarril, el pulque retoma su importancia e inicia el crecimiento de las haciendas como industrias pulqueras, tal es el caso de San Juan Tlacatecpan, San Antonio Ometusco, Zuapayuca y Venta de Cruz en el Estado de México; Santa María Tecajete, San Lorenzo, Tetlapayac, Arcos, Espejel y Chavarría en Hidalgo; San Lorenzo Soltepec, San Bartolomé del Monte e Ixtafiayuca en Tlaxcala. El mercado principal de pulque producido por estas haciendas fue el centro de México y la ciudad de Puebla, hasta el siglo XX (Monterrubio, 2007). Actualmente, el pulque es una bebida que se produce para autoconsumo, también se vende en expendios denominados “pulcatas”, se caracteriza por ser afrodisiaco y forma parte de la dieta diaria (Escalante et al., 2012; Ramírez- Rancaño, 2000).
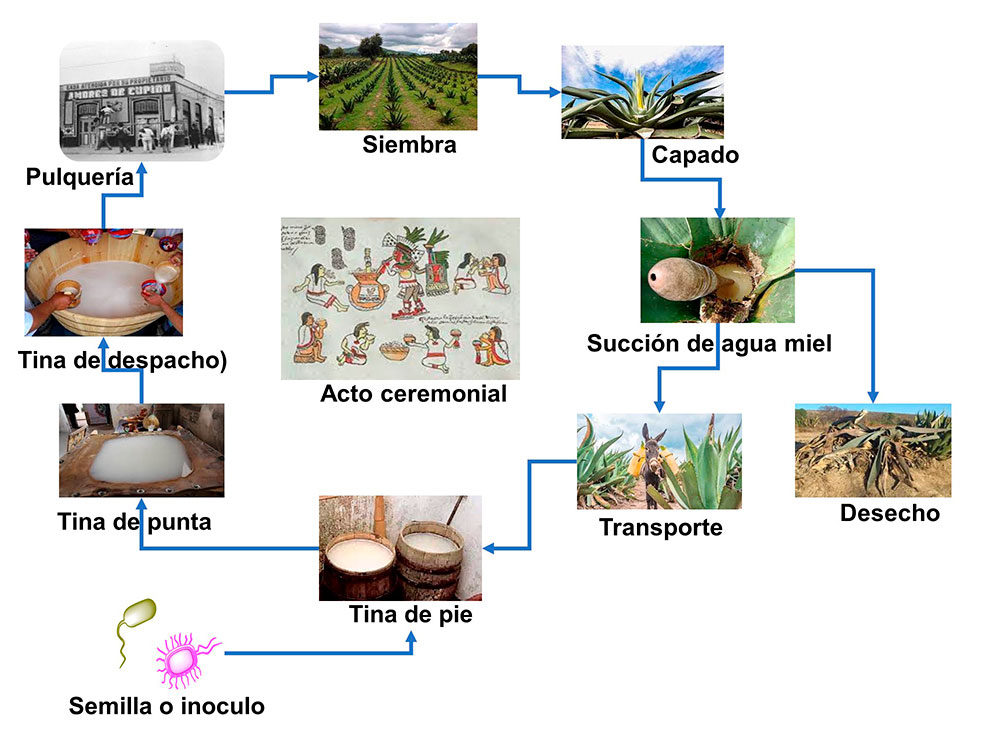
La siembra de maguey pulquero se lleva a cabo durante el cuarto creciente de la luna llena, el cuidado del cultivo es de 8 a 12 años para producir agua miel. Una vez llegado a la edad indicada se procede al capado, 4 días antes o 4 días después de luna llena, esta actividad consiste en eliminar el quiote central del maguey, se realiza una perforación en el centro, induciendo a la secreción del agua miel, posteriormente se cubre con ramas y piedras para evitar la introducción de animales. El raspado del centro del maguey es realizado por la mañana o tarde por el tlachiquero. En náhuatl “Tlachiqui” significa raspador y “ero” es la persona que hace la acción de raspar. Una vez raspada la corteza de la piña, inicia la exudación del aguamiel y su almacenamiento en el orificio, por lo que, se procede a su extracción a través de un acocote; se extrae de 3 a 4 L/día durante 3 a 4 meses y posteriormente inicia su disminución hasta secarse el maguey. El transporte del aguamiel colectado al lugar de proceso es mediante un burro o a pie.
Para iniciar la fermentación del aguamiel y producir el pulque se adiciona pulque fermentado, llamado inoculo o semilla, compuesto por un consorcio de bacterias como Zimomonas mobilis, Leuconostoc, Lactobacillus, Lactococcus, Acinetobacter y S. cerevisiae (Chacón-Vargas et al., 2020). Una vez iniciado el proceso fermentativo, se pasa a una tina de pie (barril de madera), donde se obtiene de 2 a 3 grados de alcohol y luego se traslada a la tina de la punta (bateas de madera) durante 15 días; posteriormente es llevado a la tina de despacho (tinas de madera) donde se obtiene un pulque con 7 a 8 grados de alcohol que está listo para su venta. Sin embargo, si se continúa fermentado la bebida puede alcanzar hasta 14 grados de alcohol y la bacteria inicia una etapa de etanólisis, en la cual comienza su deceso debido a la concentración alta de alcohol. El pulque en la antigüedad se bebía mediante pencas de magueyes cortos llamadas “Xoma”, ahora se utilizan recipientes de barro. Lo recomendable para dar el primer trago de esta bebida es retenerlo debajo de la lengua unos segundos y luego ingerirlo. En la actualidad también existe el pulque curado. Esta bebida es el aprovechamiento del pulque natural cuando el tinajero observa que el pulque no está en condiciones de venderse (fermentado), se procede a combinarlo con diferentes frutas y se endulza con agua miel.
Figura 1. Proceso de elaboración del pulque.
Escalante et al. (2016) reportan que las rutas metabólicas durante el proceso fermentativo del pulque consisten en 4 etapas. En la primera etapa, se lleva a cabo una fermentación alcohólica realizada por Z. mobilis y S. cerevisiae, en la que se degradan azúcares presentes en el aguamiel como sacarosa, fructosa y glucosa. En la segunda etapa, se da inicio a una fermentación láctica mediante Leuconostoc, Lactobacillus y Lactococcus, obteniendo ácido láctico, ácido acético, acetoina, diacetilo y butanodiol. En la tercera etapa, las bacterias acéticas como acetobacter y gluconobacter causan la fermentación del ácido acético. Por último, en la cuarta etapa se activa la producción del dextrano mediante las enzimas dextransacarasas del Leuconostocmesenteroides. Estos exopolisacaridos brindan la viscosidad característica al pulque. Por tanto, las propiedades sensoriales del pulque varían con base en la etapa fermentativa del proceso. La figura 2 muestra los productos terminales (etanol, ácido láctico, ácido acético, CO2 y dextrano) de las actividades metabólicas en cada etapa de fermentación. Esto se considera como una línea basal para el desarrollo de un inóculo bien definido en la fermentación del pulque.
Figura 2. Rutas de biotransformación del pulque.
Aplicaciones
El pulque, debido a su popularidad y tradición en México, se considera parte del acervo cultural de nuestro país y representa una alternativa económica que contribuye al desarrollo sustentable de comunidades rurales. Estudios recientes han demostrado que los microorganismos aislados del aguamiel y pulque generan beneficios probióticos. Leuconostocmesenteroides NRRL B‐512F puede sintetizar exopolisacáridos llamados dextrano (Etapa 4). Este exopolisacárido es uno de los principales biopolímeros obtenidos a escala industrial aplicados a la biotecnología y medicina. Las enzimas dextransacarasas son las responsables de la síntesis de dextranos. La primera aplicación industrial de los dextranos producidos por Leuconostocmesenteroides fue reemplazar al suero sanguíneo.
Los dextranos más utilizados son de 40,000 a 100,000 Dalton (Da). Por ejemplo, Leuconostocmesenteroides NRRL B‐512F produce dextranos de 40,000, 60,000 y 70.000 Da. Además, los dextranos de alto peso molecular (mayor a 100,000 Da) aumentan la posibilidad de causar reacciones alérgicas, por lo que no pueden usarse (Naessens et al., 2005). Así también, el dextrano se ha utilizado como tamiz molecular. La compañía Pharmacia Fine Chemicals (Uppsala, Sweden) en 1959 inicio la purificación y separación de macromoléculas como proteínas, ácidos nucleicos y polisacáridos. En la actualidad, esta farmacéutica se conoce como Sephadex (Separation Pharmacia Dextran). Además, el dextrano de hierro es utilizado para afrontar la falta de hierro en personas anémicas; mientras que el dextrano sulfato se utiliza como antiviral contra el virus de la gripe y el dengue (Marijke et al., 2012; Yamada et al., 2012). Por otro lado, en la industria alimentaria el dextrano se aplica en jarabes de fructosa, helados, cremas y productos de panadería (Naessens et al., 2005). No obstante, los dextranos no son aceptados como aditivos alimentarios en Estados Unidos y Europa.
En la actualidad han incrementado los registros de patentes sobre la producción de dextrano generados por microorganismos y sus recientes aplicaciones como agentes bioactivos, probióticos en alimentos funcionales, panificación y cosméticos. La cepa P45 de Leuconostocmesenteroides muestra actividad antimicrobiana contra bacterias patógenas como Listeria monocytogenes, Salmonella Typhi, Salmonella Typhimurium y Escherichia coli, tanto en modelos in vitro como in vivo (Giles et al., 2016). Por otra parte, Torres et al., (2016) reportaron que el Lactobacillus sanfranciscensis cepa LBH1068 mejora la pérdida de peso, decrece la permeabilidad intestinal y la modulación de citoquinas en ratones. Así también, los Lactobacillus sp. y Pediococcus sp. muestran actividad antimicrobiana in vitro frente a Helicobacter pylori y Staphylococcus aureus (Cervantes et al., 2019).
En conclusión, los metabolitos secundarios y microorganismos involucrados durante la producción del pulque tienen una potencial aplicación en salud y alimentación. Durante el bioproceso de producción de pulque se desarrollan microrganismos que pueden mejorar la microflora humana al actuar como probióticos. Así también, se ha demostrado que algunos de sus metabolitos podrían ser empleados como potenciales terapéuticos para mejorar la salud. Por tanto, el pulque, más allá de ser considerado una bebida alcohólica ancestral, también puede ofrecer un área de oportunidad para nuevas investigaciones en bioprocesos aplicados a la salud y la alimentación.
Referencias
Cabrera, L. G. (1992). Diccionario de aztequismos (No. 463 C32 1992).
Cervantes-Elizarrarás, A., Cruz-Cansino, N., Ramírez-Moreno, E., Vega-Sánchez, V., Velázquez-Guadarrama, N., Zafra-Rojas, Q., et al. (2019). In vitro probiotic potential of lactic acid bacteria isolated from aguamiel and pulque and antibacterial activity against pathogens. Appl. Sci. 9:601. doi: 10.3390/app9030601
Chacón-Vargas, K., Torres, J., Giles-Gómez, M., Escalante, A., and Gibbons, J. G. (2020). Genomic profiling of bacterial and fungal communities and their predictive functionality during pulque fermentation by whole-genome shotgun sequencing. Sci. Rep. 10:15115. doi: 10.1038/s41598-020-71864-4
Escalante A., Giles-Gómez M., Esquivel Flores G., Matus Acuña V., Moreno-Terrazas R., López-Munguía A., et al. (2012). Pulque fermentation, in Handbook of Plant-Based Fermented Food and Beverage Technology, ed Hui Y. H. (Boca Raton, FL: CRC Press;), 691–706.
Escalante, A., López Soto, D. R., Velázquez Gutiérrez, J. E., Giles-Gómez, M., Bolívar, F., and López-Munguía, A. (2016). Pulque, a traditional Mexican alcoholic fermented beverage: historical, microbiological, and technical aspects. Front. Microbiol. 7:1026. doi: 10.3389/fmicb.2016.01026
García Mendoza, A. (1998). Con sabor a maguey: Guía de la colección nacional de Agaváceas y Nolináceas del Jardín Botánico, Instituto de Biología-UNAM. Universidad Nacional Autónoma de México.
Giles-Gómez, M., Sandoval García, J. G., Matus, V., Campos Quintana, I., Bolívar, F., and Escalante, A. (2016). In vitro and in vivo probiotic assessment of Leuconostoc mesenteroides P45 isolated from pulque, a Mexican traditional alcoholic beverage. Springer Plus 5:708. doi: 10.1186/s40064-016-2370-7
Lappe-Oliveras, P., Moreno-Terrazas, R., Arrizon-Gavino, J., Herrera-Suarez, T., García-Mendoza, A., & Gschaedler-Mathis, A. (2008). Yeasts associated with the production of Mexican alcoholic nondistilled and distilled Agave beverages. FEMS yeast research, 8(7), 1037-1052.
Marijke, M.F.A., Schols, D. (2012). Dengue Virus Entry as Target for Antiviral Therapy. Journal of Tropical Medicine. 2012: 628475.
Monterrubio, A. L. (2007). Las haciendas pulqueras de México (Vol. 35). Universidad Nacional Autónoma de México. UNAM
Naessens, M., Cerdobbel, A., Soetaert, W., Vandamme, E.J. (2005). Leuconostoc dextransucrase and dextran: production, properties and applications. Journal of Chemical Technology & Biotechnology. 80: 845‐860.
Ramírez-Rancaño, M. (2000). Ignacio Torres Adalid y la industria pulquera. Instituto de Investigaciones Sociales- Universidad Nacional Autónoma de México. Plaza & Valdés, México.
Torres-Maravilla, E., Lenoir, M., Mayorga-Reyes, L., Allain, T., Sokol, H., Langella, P., et al. (2016). Identification of novel anti-inflammatory probiotic strains isolated from pulque. Appl. Microbiol. Biotechnol. 100, 385–396. doi: 10.1007/s00253-015-7049-4
Yamada, H., Moriishi, E., Haredy, A.M., Takenaka, N., Mori, Y., Yamanishi, K., Okamoto, S. (2012). Influenza virus neuraminidase contributes to the dextran sulfatedependent suppressive replication of some influenza A virus strains. Antiviral Research. 96: 344‐352.
Descifrando los latidos del corazón para el diagnóstico de enfermedades cardiovasculares
El corazón humano es una maravilla de la ingeniería biológica. Bombea sangre a través del cuerpo día tras día, año tras año. Pero ¿qué ocurre cuando este órgano vital muestra signos de problemas? Aquí es donde entran en juego las señales mecánicas que el corazón emite y que pueden ser fundamentales en el diagnóstico de enfermedades cardiovasculares.
Durante varios años las enfermedades del corazón han sido la principal causa de muerte en México y son un grave problema de salud pública con un alto costo económico asociado. En 2022 y de acuerdo con el Instituto Nacional de Estadística y Geografía (INEGI), del total de muertes registradas (847 mil 716), el 23.6% se atribuyeron a enfermedades del corazón. Dentro de esta categoría, las enfermedades isquémicas, caracterizadas por la reducción del flujo sanguíneo al corazón por bloqueo parcial o total de las arterias, representaron la mayoría, con un 76.5% (152 mil 960 casos). Los trastornos hipertensivos ocuparon el segundo lugar con 14.2% (28 mil 494 casos) seguidas de las relacionadas con la circulación pulmonar y otras enfermedades del corazón, con 9.0% (17 mil 960 casos). Por ello, es importante el uso y desarrollo de técnicas no invasivas para el diagnóstico y tratamiento en estos pacientes, como la electrocardiografía que registra la actividad eléctrica del corazón mediante el electrocardiograma (ECG).
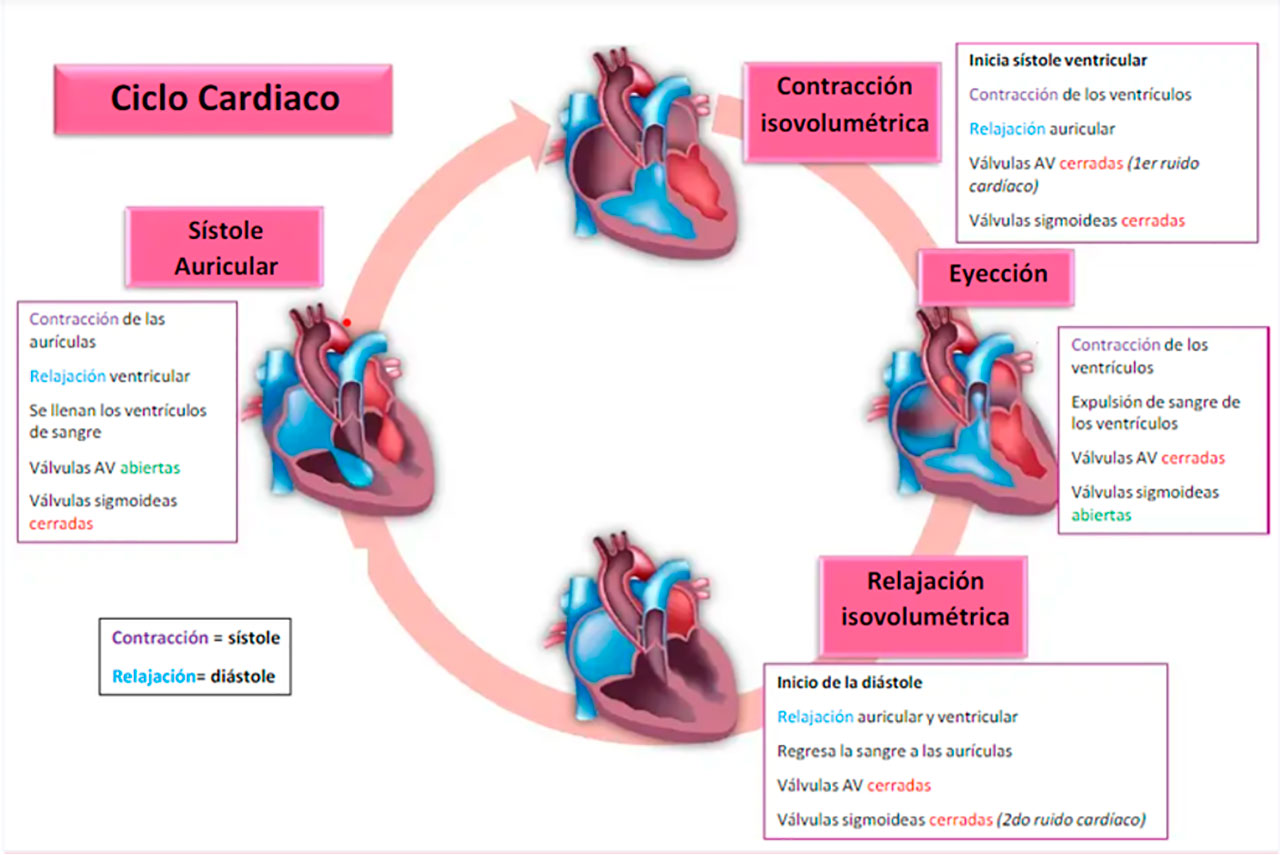
El corazón es un órgano vital del sistema cardiovascular que se encarga de bombear la sangre oxigenada e impulsar su circulación a todo el cuerpo. Para realizar estas funciones, el corazón genera continuamente impulsos eléctricos que son transmitidos por su sistema de conducción eléctrica haciendo que sus cuatro cámaras (dos aurículas y dos ventrículos) se contraigan en sucesión ordenada de tal forma que la contracción auricular (sístole auricular) va seguida de la contracción de los ventrículos (sístole ventricular) y durante la diástole todas las cámaras están relajadas. Para la conexión entre las diferentes cámaras, el corazón tiene cuatro válvulas que se abren y cierran en los instantes adecuados para asegurar que la sangre fluya solo en una dirección y evitar que retroceda (Rodrigo López, 2009). Estas válvulas se clasifican en dos grupos: auriculoventriculares (AV): válvula mitral y válvula tricúspide; y semilunares: válvula sigmoidea aórtica y válvula sigmoidea pulmonar.
Como el latido del corazón y el bombeo de la sangre generan vibraciones en el cuerpo, éstas pueden ser detectadas por transductores ubicados en su superficie, conocidos como wearables, o elementos en contacto directo con el cuerpo, como sillas, camas o básculas. Estas vibraciones han sido investigadas por más de cien años y las señales resultantes han sido denominadas de diversas maneras según el método de registro y la posición del sensor. Actualmente, el balistocardiograma (BCG), además del sismocardiograma (SCG) y el fonocardiograma (FCG) que se describirán a continuación, son las señales más consideradas en este ámbito, ya que proporcionan información de la actividad mecánica del corazón y son utilizadas en combinación con el ECG.
Sismocardiograma (SCG): vibraciones mecánicas que cuentan una historia
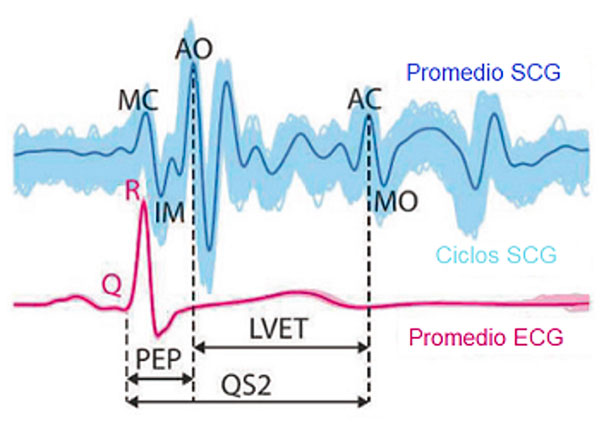
El SCG mide las vibraciones cardiacas de baja frecuencia (menores de 25 Hz) que son transmitidas a la pared del tórax, y se compone de varios puntos característicos (Figura 1): cierre de la válvula mitral (MC), apertura de la válvula aórtica (AO), inicio de la inyección rápida (RE), cierre de la válvula aórtica (AC), apertura de la válvula mitral (MO), pico del llenado rápido (RF) y sístole auricular (AS). Estos marcadores son cruciales para analizar la actividad cardíaca y evaluar su salud.
Fig. 1. Sismocardiograma (SCG) y electrocardiograma (ECG). Figura adaptada de (Sieciński & Kostka, 2017).
Se ha demostrado la utilidad del SCG en la estimación de intervalos cardíacos como el periodo de pre-eyección (PEP) y el tiempo de eyección del ventrículo izquierdo (LVET). Tanto el PEP como el LVET, son índices significativos que se utilizan ampliamente para evaluar el rendimiento del corazón (Sieciński & Kostka, 2017). El PEP representa el tiempo entre el inicio de la despolarización ventricular y el comienzo de la eyección ventricular, mientras que el LVET es un indicador de la contractilidad cardíaca y está influenciado por el sistema nervioso autónomo. Para calcular el PEP se mide desde el inicio de la onda Q del ECG hasta el pico AO del SCG (primer componente de alta velocidad). El LVET es determinado midiendo el tiempo entre los momentos de apertura y cierre de la válvula aórtica en el ciclo cardíaco.
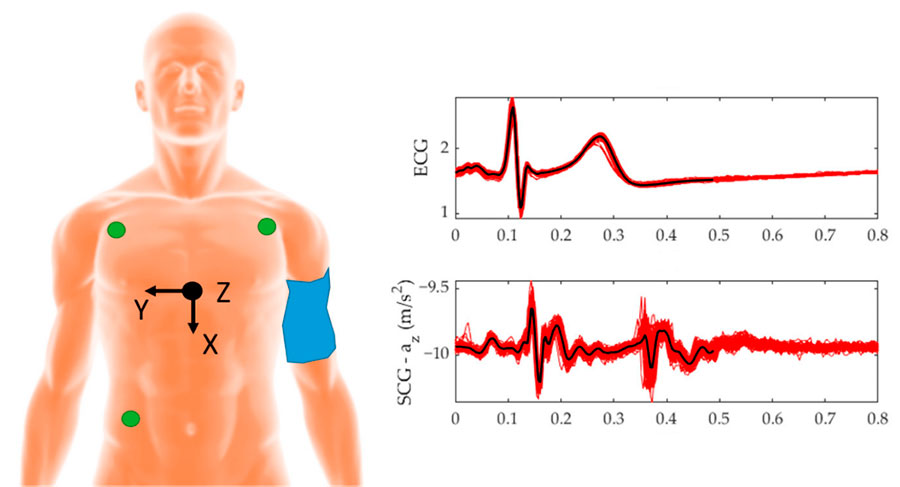
Algunos sistemas que se han desarrollado están compuestos de sensores comerciales mediante una unidad de medición inercial (IMU) colocada en la región de la apófisis xifoides del esternón (Figura 2). Esta disposición permite detectar las vibraciones y movimientos asociados con la actividad cardíaca de manera precisa. Estudios recientes demuestran que se puede detectar el SCG utilizando el acelerómetro de un teléfono móvil, un avance conocido como “m-SCG” (Landreani et al., 2018). Esto abre nuevas posibilidades para el monitoreo y diagnóstico de problemas cardiovasculares de manera más accesible y útil.
Fig. 2.Izquierda: Colocación de electrodos para ECG (verde), colocación del acelerómetro para SCG en el apófisis xifoides (negro). Derecha: ECG vs SCG (eje Z) adaptada de (D’Mello et al., 2019).
El SCG posee la capacidad de detectar algunas de las enfermedades cardiovasculares más comunes a nivel general, como la fibrilación auricular, isquemia del miocardio y hemorragia cardiaca. Esta capacidad sugiere su uso potencial como dispositivo de detección en pacientes cardiacos de alto riesgo. Un amplio estudio multicéntrico demostró que cuando se utilizaban los resultados combinados del ECG y el SCG, la precisión en la predicción de la enfermedad coronaria aumentaba de manera significativa en comparación con los resultados obtenidos solamente a partir del ECG (Wilson et al., 1993). En algunas aplicaciones, como pacientes con quemaduras, con infecciones contagiosas e incluso bebés prematuros, no resultaría factible colocar electrodos para ECG adhesivos o sensores de SCG debido a su complejidad. Por ello, se están investigando técnicas de detección del SCG sin contacto, lo que sería una mejora con respecto al SCG adquirido por sensores de contacto.
Fonocardiograma (FCG): escuchando la música del corazón
Una de las técnicas más antiguas, utilizada por el médico para el diagnóstico cardiovascular, es la auscultación, que consiste en escuchar los sonidos producidos por los órganos y vasos del cuerpo por medio del estetoscopio. El estetoscopio permite establecer un compartimiento de aire cerrado entre una región del tórax de la persona y el oído del médico. Sin embargo, este método es subjetivo, ya que depende totalmente de la capacidad de audición del médico. Aunque la auscultación todavía es el principal método para la detección y análisis de los sonidos cardíacos, existe otra técnica conocida como la fonocardiografía que utiliza un micrófono como transductor de estos sonidos integrado en un estetoscopio que elimina la interpretación subjetiva del médico.
Esta técnica registra en forma gráfica los sonidos cardíacos producidos por vibraciones de alta frecuencia (mayores de 25 Hz) generados por el corazón y por los grandes vasos sanguíneos, debido a la apertura y cierre de las válvulas cardiacas, así como del flujo de la sangre; por ello es utilizada en el diagnóstico de enfermedades valvulares. Estos sonidos se clasifican en dos categorías: ruidos y soplos cardiacos (Vermarien, 2006). Como una evolución de la tecnología de la auscultación cardiaca, estos registros se adquieren en sincronía con el ECG para comparar las relaciones temporales entre estos sonidos y los eventos mecánicos y eléctricos del ciclo cardíaco (Wen et al. 2014).
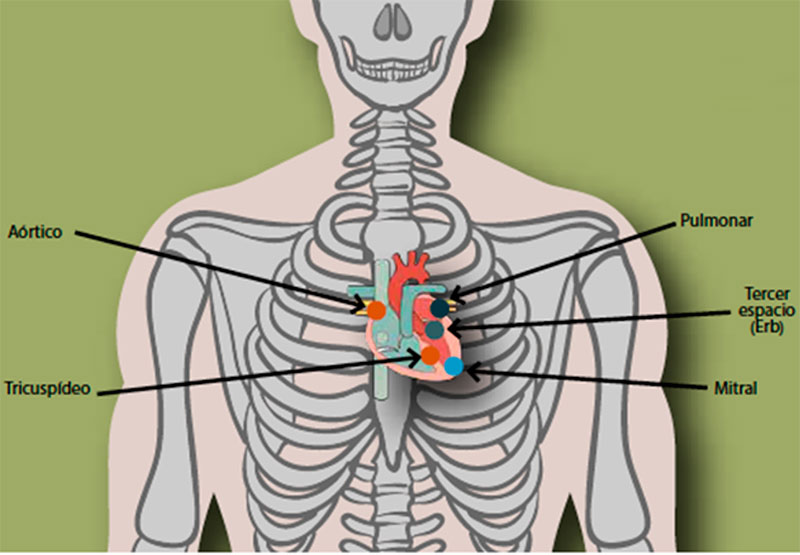
Para detectar el epicentro de los principales ruidos cardiacos existen cinco focos principales de auscultación del tórax: aórtico, pulmonar, aórtico accesorio o de Erb, tricúspideo y mitral (Figura 3).
Fig. 3. Focos de auscultación cardíaca y su ubicación anatómica. Extraída de: (Cruz-Ortega & Calderón- Monter, 2016).
La Figura 4 muestra cómo se producen los ruidos S1 y S2 durante el ciclo cardíaco debido a la coordinación de la contracción y relajación de las aurículas y ventrículos, la apertura y cierre de las 4 válvulas cardíacas y el flujo de sangre. Estos ruidos se escuchan en situaciones normales durante la auscultación con el estetoscopio.
Fig. 4. Etapas del ciclo cardíaco. Extraída de: (Tamborini, 2021).
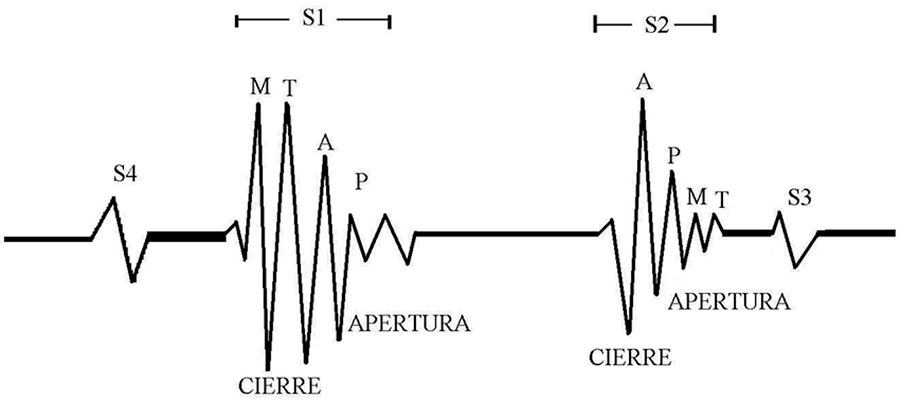
La figura 5 muestra los tipos de ruidos cardiacos y su origen. S1 y S2 marcan el inicio y el final, respectivamente de las fases del ciclo cardíaco, sístole y diástole son sonidos de alta frecuencia, mientras que S3 y S4 son de baja frecuencia que pueden escucharse en diversas afecciones. S3 indica el llenado rápido de ventrículos (diástole temprana), y está asociado a una dilatación ventricular (insuficiencia cardíaca congestiva). S4 representa el llenado tardío de los ventrículos por contracción auricular (diástole tardía) y está asociado a ventrículos no distensibles o rígidos. Las letras M (mitral), T (tricúspide), A (aortica) y P (pulmonar) señalan el cierre y la apertura de las válvulas. Estos ruidos son vibraciones o sonidos causados por la aceleración y desaceleración del flujo sanguíneo.
Fig. 5. Ruidos cardíacos y su origen. Extraída de: (Cerquera Soacha, 2005).
En este examen clínico no solo se escuchan los ruidos cardiacos, también pueden oírse los pulmones e incluso el sonido que produce el flujo sanguíneo de los vasos. La distinción entre el FCG y los sonidos pulmonares se basa en la ubicación de la auscultación, los tipos de sonidos, la relación con el ciclo cardíaco o respiratorio, la instrumentación utilizada y las fuentes de sonido. La auscultación cuidadosa y la comprensión de las características específicas de cada uno ayudarán en la diferenciación efectiva.
Con respecto a los soplos cardiacos, se trata de vibraciones o sonidos debido a la turbulencia del flujo sanguíneo, el cual se produce por el cambio de un flujo laminar a un flujo turbulento. Esto indica una disminución del área valvular (estenosis) o que la válvula no cierra correctamente y ocasiona reflujo (regurgitación o insuficiencia valvular). Los soplos son más prolongados que los ruidos cardíacos; son agudos y ásperos, a menudo contienen 2 o 3 componentes separados y pueden variar su posición corporal; pueden ser sistólicos, diastólicos o continuos. Pueden clasificarse en función de su intensidad, localización y ubicación respecto al momento del ciclo cardíaco. En un paciente con taquicardia pueden ser casi continuos. Los soplos se clasifican en intensidad en una escala de 1 al 6 (Tabla 1).
Tabla 1. Grado de intensidad de soplos cardíacos (Fonseca-Sánchez et al., 2015).
En conclusión, el SCG y el FCG se han revelado como herramientas poderosas en la detección temprana y el monitoreo de enfermedades cardiovasculares en combinación con el ECG. La capacidad de detectar sutiles alteraciones en la mecánica cardíaca, incluso antes de que se manifiesten los síntomas clínicos, brinda una oportunidad sin precedentes para una intervención temprana y un tratamiento más efectivo. Si bien aún existen desafíos en términos de estandarización y disponibilidad generalizada, el SCG y el FCG prometen una perspectiva útil en el diagnóstico y el tratamiento de enfermedades cardiovasculares, mejorando así la calidad de vida y la salud de la población.
Referencias
Rodrigo López, J. L. (2009). Válvulas cardíacas: funcionamiento y enfermedades. En A. López Farré, & C. Macaya Miguel, Libro de la salud cardiovascular del Hospital Clínico San Carlos y de la Fundación BBVA (págs. 457-462). Bilbao: Fundación BBVA.
Sieciński, S., & Kostka, P. (2017). Determining Heart Rate Beat-to-Beat from Smartphone Seismocardiograms: Preliminary Studies. Advances in Intelligent Systems and Computing, 133–140. doi:10.1007/978-3-319-70063-2_15
Landreani, F., Caiani, E. G., Golier, D., Hossein, A., Rabineau, J., Migeotte, P., & Van de Borne, P. (2018). Heartbeat Detection Using Three-Axial Seismocardiogram Acquired by Mobile Phone. Computing In Cardiology. https://doi.org/10.22489/cinc.2018.215
D’Mello, Y., Skoric, J., Xu, S., Roche, P. J. R., Lortie, M., Gagnon, S., & Plant, D. V. (2019). Real-Time Cardiac Beat Detection and Heart Rate Monitoring from Combined Seismocardiography and Gyrocardiography. Sensors (Basel, Switzerland), 19(16), 3472. https://doi.org/10.3390/s19163472
Wilson, R. A., Bamrah, V. S., Lindsay, J., Jr, Schwaiger, M., & Morganroth, J. (1993). Diagnostic accuracy of seismocardiography compared with electrocardiography for the anatomic and physiologic diagnosis of coronary artery disease during exercise testing. The American journal of cardiology, 71(7), 536–545. https://doi.org/10.1016/0002-9149(93)90508-a
Vermarien, H. (2006). Phonocardiography. En J. G. Webster, Encyclopedia of Medical Devices and Instrumentation (págs. 278-290). Hoboken, New Jersey: John Wiley & Sons.
Wen, Y.-N., Pui-Wai Lee, A., Fang, F., Jin, C.-N., & Yu, C.-M. (2014). Beyond auscultation: Acoustic cardiography in clinical practice. International Journal of Cardiology, 172(3), 548-560. doi:10.1016/j.ijcard.2013.12.298
Cerquera Soacha, E. (2005). Caracterización de estados funcionales en fonocardiografía empleando análisis acústico y técnicas de dinámica no lineal. Tesis de Maestría, Universidad Nacional de Colombia Facultad de Ingeniería y Arquitectura, Departamento de Ingeniería Eléctrica, Electrónica y Computación, Manizales. Recuperado el 16 de Abril de 2024, de https://repositorio.unal.edu.co/handle/unal/3482
El misterio del cáncer cervical: la influencia epigenética en la progresión tumoral
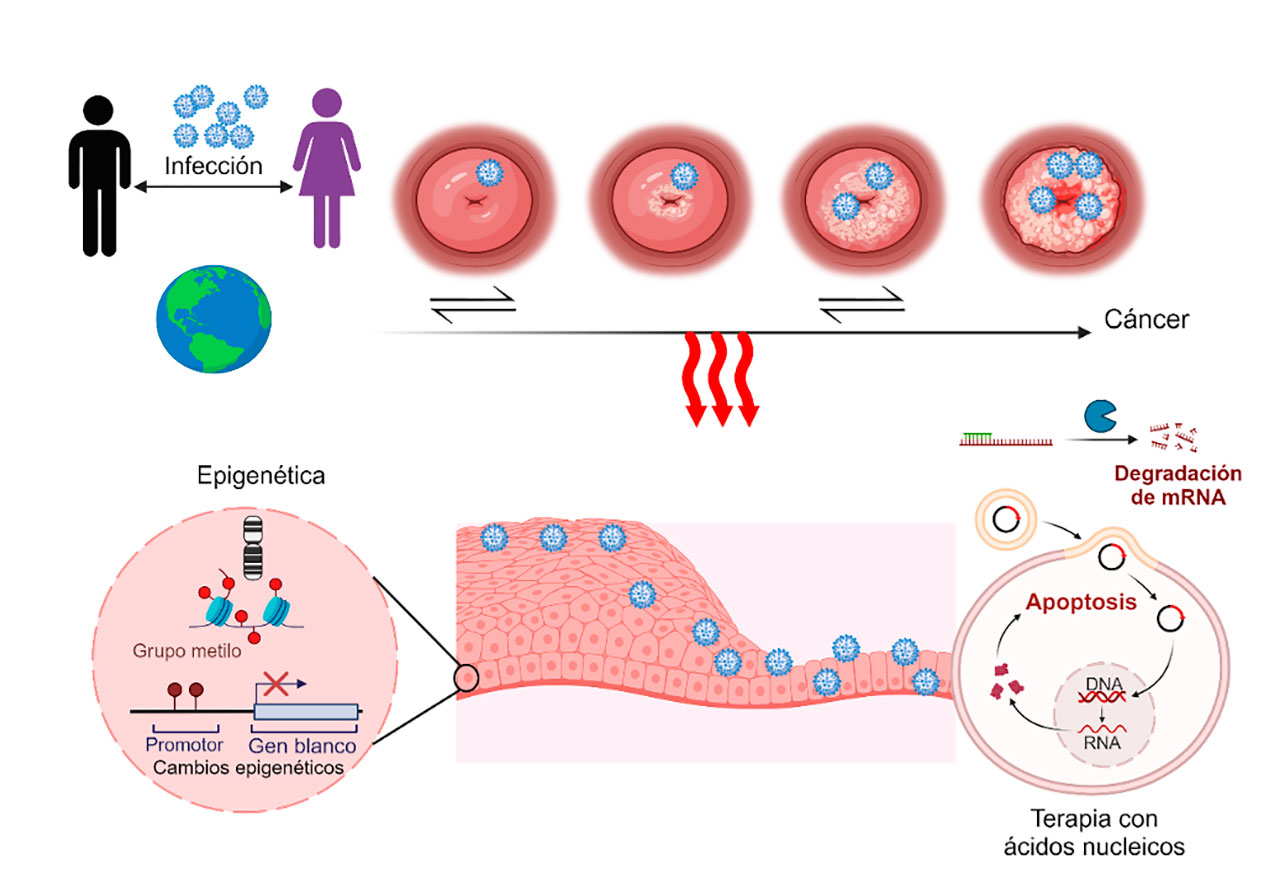
El cáncer cervical (CC) o cáncer del cuello uterino, es uno de los tipos más comunes de cáncer en mujeres en todo el mundo; su prevención y tratamiento continúan como una prioridad en la salud pública nacional y mundial. Con el tiempo, el CC ha sido objeto de una intensa inspección, debido a su naturaleza enigmática y su asociación estrecha con el virus del papiloma humano (VPH). Aunque este virus se considera como el factor de riesgo más significativo, sólo una minoría de las personas infectadas por VPH desarrolla CC, lo que sugiere la presencia de otros detonantes moleculares. En este artículo destacamos el papel crucial de los mecanismos epigenéticos en la progresión del CC, los cuales han emergido como actores clave en la desregulación de genes asociados con el cáncer. El conocimiento creciente en la epigenética del cáncer arroja información sobre nuevos enfoques para combatir esta enfermedad; es preciso resaltar que la terapia de oligonucleótidos es una estrategia potencial en el tratamiento del CC (Figura 1).
Figura 1. Esquema representativo de la infección por VPH. Progresión del cáncer cervical, participación de la epigenética, y cómo la terapia de ácidos nucleicos tiene potencial en el tratamiento del cáncer, imagen creada con BioRender.com.
La llegada de un enemigo silencioso: el virus VPH
El CC se origina comúnmente en el límite del endocérvix y el exocérvix en el cuello del útero; los cambios pre-malignos en las células escamosas del epitelio cervical parten de una infección por virus VPH, que implica 4 fases principales: (1) infección, (2) persistencia, (3) lesiones precancerosas y (4) cáncer invasivo (Yadav et al., 2023).
Los VPH son una extensa familia de más de 200 tipos de virus pequeños con un genoma de ADN que infectan epitelios y mucosas, de los cuales, los genotipos de alto riesgo incluyen a los VPH 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58 y 59 debido a su fuerte asociación con el CC. Los genotipos 16 y 18 se encuentran asociados con alrededor del 70% de tumores cervicales en el mundo (Viveros-Carreño et al., 2023; Wilailak et al., 2021). El genotipo VPH 16 es el más detectado en hombres (León et al., 2016; López Diez et al., 2017), en tanto que las cepas de VPH que más comúnmente infectan a mujeres son el VPH 16, 18, 31, 52 y 53 (Viveros-Carreño et al., 2023; Wilailak et al., 2021). No obstante, en México se ha reportado que el VPH 31 y 58 son también prevalentes (Campos et al., 2019; González-Losa et al., 2004; Rangel et al., 2020).
Los VPH de alto riesgo expresan proteínas que producen y mantienen, de manera temporal, una alta proliferación de sus células hospederas, al tiempo que nulifican la capacidad de la célula para preservar su integridad genómica, lo que tiene como consecuencia la producción de más VPH. Sin embargo, en algunas ocasiones, el genoma del VPH se integra irreversiblemente al genoma de la célula hospedera, lo que lleva a una proliferación indefinida y anómala que en algunos casos conduce a una transformación maligna (Bañuelos-Villegas et al., 2021).
El CC es diagnosticado con frecuencia, y es la causa principal de muerte por cáncer en las mujeres de países subdesarrollados. Según datos reportados por el GLOBOCAN, se estiman 604 000 casos nuevos y 342 000 muertes en todo el mundo hasta 2020 (Sung et al., 2021).
La presencia del VPH en las poblaciones latinoamericanas es alta: 12.3% en América del Sur y 20.4% en América Central. Esta población muestra alta presencia de VPH en una edad menor a 35 años (Scott-Wittenborn y Fakhry 2021). Adicionalmente, en México el CC ocupa el segundo lugar entre los tumores que afectan a la mujer, con una prevalencia de 16.1%, incidencia del 15.9% y una mortalidad del 13.7% (Wilailak et al., 2021; Yadav et al., 2023). El 99.7% de casos de CC se relacionan con la infección por el VPH de alto riesgo; ésta se puede transmitir no sólo por relaciones sexuales con penetración, sino también por contacto genital piel con piel. Aunque la mayoría de las infecciones por VPH se resuelve espontáneamente dentro de 12-24 meses, la infección persistente puede causar CC en las mujeres en un plazo entre 10 y 15 años (Wilailak et al., 2021; Yadav et al., 2023).
Se sabe que los hombres o parejas sexuales de mujeres con lesiones precursoras de alto grado, constituyen un importante reservorio y vector de infección por VPH (López Diez et al., 2017). Otros factores del CC son el inicio temprano de la actividad sexual, múltiples parejas sexuales, antecedentes de infecciones de transmisión sexual como VIH, historial de lesiones vulvares o vaginales, tabaquismo, píldoras anticonceptivas orales, multiparidad, entre otras (Wilailak et al., 2021).
El CC es una de las formas de cáncer que se ha tratado con más éxito, siempre que se detecte oportunamente y se trate de forma eficaz. En 2020, la Organización Mundial de la Salud (OMS) adoptó una estrategia global para la eliminación del CC a través de una intervención triple (Wilailak et al., 2021). Este control integral del CC incluye:
La prevención primaria mediante la vacunación contra el VPH, es uno de los principales desafíos en los países de ingresos bajos. Se busca que el 90% de las niñas estén completamente vacunadas a la edad de 15 años. Actualmente la OMS precalifica tres vacunas profilácticas contra el VPH: la vacuna bivalente Cervarix™ (GlaxoSmithKline, Rixensart, Belgium), la tetravalente Gardasil™ (Merck, West Point, PA, USA) y la vacuna nonavalente Gardasil-9™ (Merck) (Yousefi et al., 2022).
La prevención secundaria pretende que el 70% de las mujeres sean examinadas a la edad de 35 años y nuevamente a los 45 años, incluidos un programa de detección de alta cobertura, la promoción de la salud y el tratamiento precanceroso temprano, especialmente en mujeres no vacunadas y mujeres infectadas con subtipos distintos del VPH 16 y el VPH 18 (Viveros-Carreño et al., 2023; Wilailak et al., 2021).
El objetivo de la prevención terciaria es mejorar la calidad de vida de la paciente y reducir la discapacidad mediante un tratamiento y rehabilitación eficaces (Viveros-Carreño et al., 2023; Wilailak et al., 2021).
Es significativo resaltar que a las mujeres mayores de 15 años, que nunca han estado expuestas al VPH, se les recomienda recibir tres dosis de vacunación para la prevención del CC. Es transcendental implementar programas de vacunación y educación sexual para niñas y niños con el fin de aumentar su conciencia sobre las enfermedades sexualmente transmitidas (EST), fomentando salud sexual, reduciendo las conductas sexuales de alto riesgo, promoviendo el uso de preservativos, el abandono del hábito de fumar y la revisión ginecológica. Estas intervenciones deben adaptarse, según la población objetivo, la edad y la cultura.
La epigenética y el CC
Las modificaciones epigenéticas son cambios que ocurren en el material genético (ADN) no asociadas a cambios en la secuencia y que son parte de un proceso evolutivo natural. La evidencia sugiere que las alteraciones en los patrones de expresión génica son gobernadas por modificaciones epigenéticas que pueden resultar en cáncer. Estas alteraciones ocurren a través de cuatro mecanismos principales denominados: metilación del ADN, modificación de histonas, regulación de ARN no codificante, y regulación de cromatina (Liu et al., 2023). A continuación, se describe cada aspecto desde una perspectiva simple y general (Albulescu et al., 2021; Liu et al., 2023; Yadav et al., 2023).
-Metilación del ADN: es el mecanismo central de las anomalías epigenéticas de los tumores; implica la transferencia de un grupo metilo bajo la acción de enzimas llamadas ADN metiltransferasas. En el CC, el nivel de metilación del ADN se modifica, promoviendo con esto la regulación de múltiples genes, induciendo un efecto fenotípico en las células infectadas después de integración del ADN del VPH.
-Modificación de proteínas llamadas histonas: son cruciales en la regulación y pueden ser categorizadas por la alteración de grupos químicos pequeños o macroscópicos. Por ejemplo, la acetilación de histonas produce un cambio espacial de una estructura llamada nucleosoma, la unidad básica de la cromatina, al inducir pérdida de la estructura de la cromatina, y con ello, regulando la trascripción génica.
-Moléculas de ARN no codificante (ARNnc): no codifican a proteínas, pero juegan un papel importante en la regulación epigenética. En correspondencia con su tamaño y sus características estructurales, ARNnc puede ser clasificado en diferentes grupos. Recientemente se ha identificado a los ARNnc como reguladores biológicos importantes, tanto de manera normal como en las enfermedades.
-Regulación de la cromatina: esta estructura determina las propiedades celulares y su homeostasis. En células cancerosas existe una alteración de esta estructura, promoviendo una proliferación anormal (Albulescu et al., 2021; Liu et al., 2023).
¿Qué tiene que ver la epigenética con el CC?
Los marcadores epigenéticos asociados con eventos moleculares inducidos por el VPH se pueden utilizar para clasificar a las mujeres con riesgo de desarrollar cáncer de cuello uterino, la respuesta a la terapia o el pronóstico de la enfermedad (Albulescu et al., 2021; Da Silva et al., 2021).
El ADN es como un libro de instrucciones, y la epigenética como notas en los márgenes que regulan qué partes se leen. En el CC, cambios epigenéticos pueden apagar genes que controlan el crecimiento celular normal. Es como desordenar las instrucciones y hacer que la célula afectada se comporte de manera anormal. Los factores ambientales, como infecciones por el virus VPH, pueden influir en estos cambios. Comprender estas “notas” epigenéticas nos acerca a tratamientos personalizados y abre la puerta a prevenir o tratar el CC de manera más efectiva. ¿Cómo sería posible establecer una estrategia de tratamiento a través de la epigenética? Utilizando secuencias pequeñas de ácidos nucleicos que puedan regular o inhibir el cáncer; aquí es donde intervienen el ARNnc y los oligonucleótidos como tratamiento.
Terapia de oligonucleótidos: ¿una nueva esperanza?
La quimioterapia convencional para el cáncer utiliza agentes cuyo efecto terapéutico es por la inducción de la muerte celular; sin embargo, se puede desarrollar resistencia a esta terapia. Por su parte, los fármacos desarrollados recientemente, conocidos como “terapia dirigida”, pueden mostrar menor toxicidad no deseada, pero son generalmente menos efectivos. En consecuencia, existe la necesidad urgente de desarrollar nuevos fármacos eficaces. Una estrategia emergente consiste en centrarse en procesos moleculares finos, mediante el uso de la terapia con oligonucleótidos (De Rosa et al., 2010).
Los oligonucleótidos son fragmentos cortos de ácidos nucleicos sintéticos, capaces de corregir selectivamente la expresión de proteínas en las células. Existen varios tipos, cada uno con un mecanismo de acción diferente, ofrecen un nuevo y prometedor concepto de terapia contra el cáncer. Las modificaciones químicas en la estructura principal del oligonucleótido pueden mejorar su función y efecto. Adicionalmente, se conocen desde hace tiempo estrategias prometedoras o sistemas para su administración eficiente (Alvarez-Salas y DiPaolo, 2007; DiPaolo y Alvarez-Salas, 2004).
Conclusión
Los mecanismos epigenéticos juegan un papel importante en la progresión del CC. Si bien el VPH ha sido identificado como un factor de riesgo decisivo, otros detonantes moleculares también desempeñan un papel significativo para el desarrollo de la enfermedad. El entendimiento de esta compleja red de interacciones moleculares es fundamental para desarrollar enfoques terapéuticos más precisos enfocados en la terapéutica de oligonucleótidos, y estrategias de prevención efectivas. Estos avances prometedores en la comprensión de la epigenética del CC, abren nuevas posibilidades para combatir esta enfermedad presente en todo el mundo.
Referencias
Albulescu, A., Plesa, A., Fudulu, A., Iancu, I. V., Anton, G., & Botezatu, A. (2021). Epigenetic approaches for cervical neoplasia screening (Review). Experimental and Therapeutic Medicine, 22(6), 1481. https://doi.org/10.3892/etm.2021.10916
Alvarez-Salas, L., & DiPaolo, J. (2007). Molecular Approaches to Cervical Cancer Therapy. Current Drug Discovery Technologies, 4, 208–219. https://doi.org/10.2174/157016307782109661
Bañuelos-Villegas, E. G., Pérez-yPérez, M. F., & Alvarez-Salas, L. M. (2021). Cervical Cancer, Papillomavirus, and miRNA Dysfunction. Frontiers in Molecular Biosciences, 8, 758337. https://doi.org/10.3389/fmolb.2021.758337
Campos, R. G., Malacara Rosas, A., Gutiérrez Santillán, E., Delgado Gutiérrez, M., Torres Orozco, R. E., García Martínez, E. D., Torres Bernal, L. F., & Rosas Cabral, A. (2019). Unusual prevalence of high-risk genotypes of human papillomavirus in a group of women with neoplastic lesions and cervical cancer from Central Mexico. PLOS ONE, 14(4), e0215222-. https://doi.org/10.1371/journal.pone.0215222
Da Silva, M. L. R., De Albuquerque, B. H. D. R., Allyrio, T. A. D. M. F., De Almeida, V. D., Cobucci, R. N. D. O., Bezerra, F. L., Andrade, V. S., Lanza, D. C. F., De Azevedo, J. C. V., De Araújo, J. M. G., & Fernandes, J. V. (2021). The role of HPV-induced epigenetic changes in cervical carcinogenesis (Review). Biomedical Reports, 15(1), 60. https://doi.org/10.3892/br.2021.1436
De Rosa, G., De Stefano, D., & Galeone, A. (2010). Oligonucleotide delivery in cancer therapy. Expert Opinion on Drug Delivery, 7(11), 1263–1278. https://doi.org/10.1517/17425247.2010.527942
DiPaolo, J. A., & Alvarez-Salas, L. M. (2004). Advances in the development of therapeutic nucleic acids against cervical cancer. Expert Opinion on Biological Therapy, 4(8), 1251–1264. https://doi.org/10.1517/14712598.4.8.1251
González-Losa, M. del R., Rosado-Lopez, I., Valdez-González, N., & Puerto-Solı́s, M. (2004). High prevalence of human papillomavirus type 58 in Mexican colposcopy patients. Journal of Clinical Virology, 29(3), 203–206. https://doi.org/https://doi.org/10.1016/S1386-6532(03)00138-0
León, D., Retamal, J., Silva, R., Mieville, C. I. S., Guzmán, P., Briceño, G., & Brebi, P. (2016). Detección molecular de agentes infecciosos de transmisión sexual en un grupo de hombres sintomáticos y su relación con la conducta sexual. Revista Chilena de Infectología, 33, 505–512.
Liu, H., Ma, H., Li, Y., & Zhao, H. (2023). Advances in epigenetic modifications and cervical cancer research. Biochimica et Biophysica Acta (BBA) – Reviews on Cancer, 1878(3), 188894. https://doi.org/https://doi.org/10.1016/j.bbcan.2023.188894
López Diez, E., Pérez, S., Iñarrea, A., de la Orden, A., Castro, M., Almuster, S., Tortolero, L., Rodríguez, M., Montero, R., & Ojea, A. (2017). Prevalence and concordance of high-risk papillomavirus infection in male sexual partners of women diagnosed with high grade cervical lesions. Enfermedades Infecciosas y Microbiología Clínica, 35(5), 273–277. https://doi.org/https://doi.org/10.1016/j.eimc.2016.02.014
Rangel, S., Basilio, H.-S., Thompson-Bonilla, M., Hector, R.-G., & Alvarez-Salas, L. (2020). A Pilot Study on the HPV Type Frequency in a Federal High-Specialty Hospital of Mexico City: Is HPV16 Our Main Problem? SN Comprehensive Clinical Medicine, 2. https://doi.org/10.1007/s42399-020-00245-9
Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., & Bray, F. (2021). Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer Journal for Clinicians, 71(3), 209–249. https://doi.org/https://doi.org/10.3322/caac.21660
Viveros-Carreño, D., Fernandes, A., & Pareja, R. (2023). Updates on cervical cancer prevention. International Journal of Gynecologic Cancer, 33(3), 394. https://doi.org/10.1136/ijgc-2022-003703
Wilailak, S., Kengsakul, M., & Kehoe, S. (2021). Worldwide initiatives to eliminate cervical cancer. International Journal of Gynaecology and Obstetrics: The Official Organ of the International Federation of Gynaecology and Obstetrics, 155 Suppl 1(Suppl 1), 102–106. https://doi.org/10.1002/ijgo.13879
Yadav, C., Yadav, R., Chabbra, R., Nanda, S., Ranga, S., Kadian, L., & Ahuja, P. (2023). Overview of genetic and epigenetic regulation of human papillomavirus and apoptosis in cervical cancer. Apoptosis, 28(5), 683–701. https://doi.org/10.1007/s10495-023-01812-w
Yousefi, Z., Aria, H., Ghaedrahmati, F., Bakhtiari, T., Azizi, M., Bastan, R., Hosseini, R., & Eskandari, N. (2022). An Update on Human Papilloma Virus Vaccines: History, Types, Protection, and Efficacy. Frontiers in Immunology, 12, 805695. https://doi.org/10.3389/fimmu.2021.805695
Sistema de transporte nuclear de las importinas alfa y beta en Entamoeba histolytica
Introducción
Entamoeba histolytica es un parásito protozoario perteneciente al filo amoebozoa y es conocido por ser el agente etiológico de la disentería amebiana, conocida como amebiasis en humanos. De manera silvestre E. histolytica experimenta un ciclo de vida complejo que involucra a un hospedero (Figura 1). El 90% de los pacientes infectados presentan síntomas y pueden mantener los parásitos en el lumen del colon, mientras que el 10% restante no muestra síntomas (Begum et al., 2021). En el mundo, aproximadamente 50 millones de personas son infectadas anualmente, con más de 100,000 muertes debido a la disentería amebiana (Zulfiqar et al., 2023). En 2022 en México se reportaron más de 162,000 casos de amebiasis intestinal y 428 casos de abscesos hepáticos causados por la infección con este parásito protozoario (Revista Epidemiológica Nacional).
Ciclo de vida de E. histolytica
Los quistes y los trofozoítos son transmitidos principalmente por la materia fecal. Los quistes suelen estar presentes en las heces sólidas mientras que los trofozoítos están en el material diarreico. La infección con E. histolytica ocurre principalmente mediante la ingestión de alimentos o agua contaminada con quistes maduros (Marie and Petri, 2013; Ralston, 2015). También se han reportado infecciones mediante contacto sexual oro-anal. Una vez dentro del huésped, el enquistamiento se lleva a cabo en el intestino delgado y los trofozoítos son liberados para posteriormente migrar al intestino grueso. Se puede presentar una infección no invasiva en la cual los trofozoítos permanecen en el lumen intestinal sin que el huésped presente algún tipo de sintomatología. Los trofozoítos se dividen por fisión binaria y son capaces de infectar la mucosa intestinal, los vasos sanguíneos y sitios extraintestinales como cerebro, pulmones e hígado. Los quistes están adaptados para sobrevivir de días a semanas y permanecer en un estado infeccioso debido a que sus paredes celulares le confieren resistencia al medio externo. De manera contrastante, los trofozoítos no resisten al medio exterior una vez que son expulsados del huésped, y si son ingeridos oralmente, pueden no resistir a los ácidos gástricos producidos en la digestión estomacal. Los síntomas clínicos incluyen dolor abdominal, pérdida de peso, diarrea o inclusive diarrea con sangre, que puede durar varias semanas. Una infección sistémica puede perdurar e incluso años después de la infección inicial, en la que el órgano comúnmente más afectado es el hígado en el que se forman abscesos hepáticos; también se puede presentar ictericia, fiebre, tos, escalofríos, aumento en el volumen del hígado y pérdida de peso (Haque et al., 2003; Pritt and Clark, 2008).
Patogenicidad
Los mecanismos de patogenicidad de E. histolytica incluyen adherencia, citotoxicidad y fagocitosis/trogocitosis. La adherencia es un proceso fundamental para la infección de E. histolytica. La secreción de cistein-proteasas ocasiona el adelgazamiento de las mucinas del colon del huésped y permite a los trofozoítos unirse a las capas de mucina por medio de la vía de Gal/lectina. Posteriormente, los trofozoítos son capaces de unirse a las células epiteliales del huésped (Begum et al., 2015; Singh et al., 2016).
Después de la adherencia, se puede entrar a un proceso en el que E. histolytica mata a las células del huésped y a las bacterias intestinales, haciendo interacción directa entre el parásito con las células del huésped y la secreción de proteinasas para facilitarle la infección que puede desatar un proceso de activación de apoptosis (Christy and Petri, 2011; Becker et al., 2010). Este parásito protozoario codifica para proteínas llamadas amebaporos que pueden formar poros en las bicapas lipídicas cuya función es la fagocitosis de material extracelular por medio de fagosomas (Berninghausen y Leippe, 1997; Ralston, 2015).
En años recientes se ha fortalecido la hipótesis de que la trogocitosis es el principal mecanismo que utiliza E. histolytica para engullir y alimentarse de células vivas (Ralston et al., 2014). Para unirse a las células vivas del huésped E. histolytica lleva a cabo un rearreglo de los filamentos de actina y posteriormente va engullendo pequeñas porciones de las células del huésped, ocasionando una muerte celular que conduce a una disociación en la interacción de E. histolytica con la célula huésped (Ralston, 2014; Ralston, 2015).
El proceso de fagocitosis es crucial para la obtención de nutrientes para E. histolytica al invadir los tejidos del huésped y al interactuar las partículas con los receptores de superficie celular del parásito. Este fenómeno se lleva a cabo debido a un rearreglo de los microfilamentos de actina que proveen la formación del fagosoma para el engullimiento del material extracelular (Christy y Petri, 2011). La actina es una proteína fundamental en los fagosomas tempranos, ya que están rodeados de filamentos de actina que se van despolimerizando gradualmente conforme el fagosoma va madurando. Esta maduración se ve controlada por proteínas pertenecientes a la superfamilia Ras, principalmente las proteínas Rab, fosfatidilinositol y proteínas cinasas (Saito-Nakano et al., 2005; Somlata et al., 2011; Somlata et al., 2012; Avalos-Padilla et al., 2015; Verma et al., 2015; Verma et al., 2016; Hanadate et al., 2016). Similar a eucariontes multicelulares, parece que este proceso se rige por el complejo ESCRT (del inglés: Endosomal Sroting Complexes Required for Transport) (Lopez-Reyes, 2010). Adicionalmente se ha visto que el Ca2+ está implicado en el proceso de fagocitosis, principalmente en la formación de fagosomas en donde han sido identificadas algunas proteínas incluidas en este proceso como son: EhCaBP3 y EhCaBP1 que se localizan de manera ubicua en el cuerpo del parásito (Sahoo et al., 2004).
Figura 1. Ciclo de vida de E. histolytica. A) Los quistes y trofozoítos son transmitidos a través de la materia fecal. B) El agua o la comida contaminada son el vehículo de la ingesta de quistes. C) El enquistamiento ocurre en el intestino. D) Los trofozoítos son liberados cuando migran al intestino grueso. E) Los trofozoítos se dividen por fisión binaria y pueden producir quistes que finalmente pasan a la materia fecal y así cerrar el ciclo (Tomado y modificado de https://www.cdc.gov/dpdx/amebiasis/index.html).
Tratamientos actuales
La invasión intestinal asintomática con E. histolytica, frecuentemente se trata con yodoquinol y paromomicina, mientras que el tejido invadido es tratado con metronidazol (Figura 2); sin embargo, el metronidazol es un antibiótico no especifico usado para combatir bacterias anaerobias e infecciones por protozoarios. Además, el metronidazol tiene efectos cardiovasculares adversos y problemas gastrointestinales (Kimura et al., 2007; Stanley, 2003). Existen pocos estudios con drogas distintas al metronidazol, extractos herbales en pacientes con amebiasis o con una droga llamada auranofin cuyo uso es para tratar la artritis reumatoide. Se sabe que auranofin es una droga cuyo blanco es la tiorredoxina reductasa que previene la reducción de tiorredoxina y potencia la sensibilidad de los trofozoítos al reaccionar con el oxígeno (Sha et al., 2016, Debnath et al., 2012). Actualmente se desarrollan moléculas que inhiben el transporte nuclear para el tratamiento de infecciones virales como el VIH y el Dengue, y se ha probado el uso de ivermectina para inhibir el trasporte nuclear de la RNA Pol I en Trypanosoma cruzi los cuales se podrían probar para la disentería amebiana; sin embargo, se requiere de la investigación farmacológica y de biología molecular en E. histolytica (Wagstaff et al., 2012; Canela-Pérez et al., 2018). En particular, se necesita del conocimiento de los sistemas de transporte de proteínas hacia y desde el núcleo para evaluar la factibilidad de uso de fármacos como la ivermectina para tratar la amibiasis.
Figura 2. Drogas para el tratamiento de la disentería amebiana. Estructuras moleculares del yodoquinol (A), de la paromomicina (B) y del metronidazol (C).
Transporte nuclear de proteínas mediado por Importina α e Importina β
Es bien sabido que la vía de importación nuclear α/β está bien conservada en eucariontes, como Saccharomyces cerevisiae, Mus musculus y Homo sapiens (Wente and Rout et al., 2010). Existe poca literatura de esta vía de transporte nuclear de proteínas en protozoarios, por lo tanto, la vía de transporte nuclear canónica α/β que involucra a la importina alfa (IMPα) y beta (IMPβ), no ha sido estudiada en E. histolytica. Sin embargo, hemos realizado una búsqueda informática y encontramos anotados los genes codificantes para estas proteínas en la base de datos AmoebaDB (números de acceso: EHI_025350 para IMPα y EHI_036520 para IMPβ).
El transporte nuclear de proteínas es un proceso altamente complejo que transcurre a través de los complejos de poro nuclear (NPC). En esta vía participan proteínas llamadas importinas y exportinas pertenecientes a la familia de las carioferinas (Strambio-De-Castillia et al., 2010). El mecanismo de translocación de proteínas al nucleoplasma mediado por IMPα e IMPβ se conoce como vía clásica de importación nuclear de proteínas (Cauntain et al., 2015). Las proteínas nucleares son transportadas desde el citoplasma al interior de núcleo (Cargo) y generalmente tienen motivos proteínicos conocidos como señales de localización nuclear (NLS), que son reconocidos por la proteína adaptadora IMPα formando el complejo Cargo/IMPα; posteriormente la proteína receptora IMPβ se une al dominio IBB (Importin Beta Binding domain) de IMPα formando el complejo trimérico de transporte Cargo/IMPα/IMPβ (Stewart y Rhodes, 1999; Stewart, 2007). Este complejo es translocado al nucleoplasma atravesando el NPC mediado por IMPβ al interactuar con dominios ricos en fenilalanina y glicina (FG) de las nucleoporinas que conforman el NPC (Ma et al., 2016). Una vez que el complejo trimérico se encuentra en el nucleoplasma, una proteína de unión a GTP llamada Ran-GTP (perteneciente a la familia RAS) se une a IMPβ generando un cambio conformacional y así disgregando el complejo trimérico Cargo/IMPα/IMPβ (Vetter et al., 1999). IMPβ permanece unida a Ran-GTP y es exportada al citoplasma; así, Ran-GTP cumple una función regulatoria del transporte nuclear al mantener un ciclo de unión con GTP y GDP, siendo el gradiente de Ran-GTP mayoritario en el núcleo (Görlich et al., 1996; Hoelz y Blobel, 2004). La unión de GTP a Ran es modulada por el factor de intercambio de nucleótido de guanina (RanGEF) en el núcleo y la proteína activadora de GTPasa de Ran (Ran GAP), ambas proteínas distribuidas de manera asimétrica entre el núcleo y citoplasma controlan la unión de Ran a GTP o GDP y por consiguiente, la regulación del transporte nuclear (Bischoff et al., 1994; Moroianu et al., 1996; King et al.,2006). Por otra parte, IMPα es transportada al citoplasma mediante la proteina CAS (del inglés: the Celular Apoptosis Susceptibility gene product), en complejo con RanGTP (Kutay et al., 1997; Stewart, 2007). Los complejos exportados RanGTP-CAS-IMPa y RanGTP-IMPβ son disociados por la hidrólisis de RanGTP generando RanGDP en el citoplasma. La actividad de GTPasa de Ran es estimulada por la proteína activadora de GTPasa RanGAP1 (Seewald et al., 2002). Adicionalmente, las proteínas RanBP1 o RanBP2 estimulan a RanGAP1 para la activación de GTPasa de Ran (Matunis et al., 1996; Vetter et al., 1999; Ritterhoff et al., 2016). El transporte nuclear de RanGDP depende de NTF2 (Nuclear Transport Factor 2) que tiene afinidad por los dominios FG de las nucleoporinas que conforman el NPC (Stewart, 2000; Jovanovic-Talisman et al., 2009). Una vez en el nucleoplasma NTF2-RanGDP es disociado por la reconversión de RanGDP a Ran-GTP realizado por RanGEF (Bischoff y Ponstingl, 1991). Mientras que la proteína CAS puede regresar nuevamente al núcleo por sí misma. Las proteínas exportadas al citoplasma contienen un motivo llamado señal de exportación nuclear (NES) que es reconocido por factores de exportación nuclear llamados exportinas; estas son proteínas pertenecientes también a la familia de las carioferinas. El proceso de exportación nuclear requiere la participación de Ran-GTP que se une a las exportinas (Xu y Massagué, 2004) (Figura 3).
En E. histolytica, la vía de transporte nuclear clásica no ha sido estudiada; sin embargo, en la base de datos AmoebaDB identificamos los genes que codifican para las proteínas lMPα, IMPβ, CAS y Ran, RanGEF, RanBP1 y RanGAP, indicando que la vía se encuentra conservada en este parásito protozoario, similar a lo que ocurre en tripanosomatidos (Canela-Pérez et al., 2019). Se ha reportado que en los genomas de L. major y T. brucei se han identificado las proteínas NTF2, RanGAP, RanBP1 y RCC1 (Casanova et al., 2008).
Figura 3. Vía de transporte nuclear de las importinas α y β. El adaptador inicial de la vía IMPα (morado) reconoce la NLS de la proteína cargo (mostaza) y posteriormente se une IMPβ (verde) para formar el complejo trimérico de importación nuclear IMPα/IMPβ/cargo. El complejo trimétrico de importación atraviesa el complejo de poro nuclear y una vez en el nucleoplasma, la proteína Ran-GTP se une a IMPβ para disgregar el complejo trimérico de importación. La IMPβ permanece unida a Ran GTP (azul) y es exportada al citoplasma, mientras que IMPα es reconocida por la exportina RCC1 (rojo) unida a RanGTP para ser exportada al citoplasma. Las proteínas con una señal de exportación nuclear (negro) son reconocidas por la exportina RCC1 unida a Ran-GTP, y este complejo trimétrico de exportación nuclear es transportado al citoplama. Una vez en el citoplasma, las proteínas RanGAP y Ran BP1 llevan a cabo la hidrólisis de GTP por GDP liberando los complejos y dejando disponibles a IMPα, IMPβ y la proteína con NES. El reciclamiento de GTP al núcleo se lleva a cabo por el factor NTF2 (amarillo) que tiene afinidad por Ran-GDP. Una vez en el nucleoplasma la proteína Ran-GEF realiza el cambio de GDP por GTP en Ran generando la disociación de NTF2 de Ran-GTP.
Importina α
IMPα es una proteína perteneciente a la familia de las carioferinas que participa como adaptador en la vía clásica de importación nuclear de proteínas, al reconocer las cNLS de las proteínas cargo (Goldfarb et al., 2004; Oka y Yoneda, 2018). En su extremo amino muestra el dominio IBB y en su extremo carboxilo presenta 10 dominios Armadillo (ARM) (Cingolani et al., 1999; Kobe, 1999). Los dominios ARM fueron descritos en D. melanogaster en la proteína Armadillo y se conforman por tres hélices a (H1, H2 y H3) (Riggleman et al.,1989). Los dominios ARM generan una estructura conocida como alfa solenoide constituida por motivos hélice-giro-hélice (Fournier et al., 2013) que giran consecutivamente en la estructura de IMPα creando un surco a lo largo del eje de la superhélice de la proteína en donde se localizan los sitios de unión a las cNLS, específicamente los motivos conservados (^R/K) XXWXXXN), en donde ^R/K es cualquier residuo a excepción de Arg o Lys (principalmente residuos ácidos o hidrofóbicos), mientras que X es cualquier aminoácido (Conti et al., 1998; Conti y Kuriyan, 2000). Los residuos encargados de la interacción con la cNLS son Asn y Trp que generan interacción con Arg y Lys (Fontes et al., 2000). Las cNLS monopartitas y bipartitas se unen a los sitios de unión de IMPα en una conformación extendida (Fontes etal., 2003). Mediante análisis estructurales se ha descrito que las cNLSs monopartitas se unen al bolsillo mayor de IMPα, mientras que las cNLS bipartitas se unen a los bolsillos menor y mayor (Yang et al., 2010). Por otra parte, las mutaciones en residuos en ambos bolsillos de manera simultanea interrumpen la unión de NLS bipartitas (Leung et al., 2003). A la fecha, se ha logrado determinar la estructura de IMPα de H. sapiens (Dias et al., 2009), S. cerevisiae (Conti y Kuriyan, 2000), M. musculus (Kobe, 1999; Fontes et al., 2000), Oryza sativa (Chang et al., 2012; Chang et al., 2014), Arabidopsis thaliana (Wirthmueller et al., 2015) y Neurospora crassa (Bernardes et al., 2015).
IMPα no ha sido estudiada en E. histolytica, sin embargo, la hemos modelado usando el programa alphafold en alphafold.ebi.ac.uk y buscamos sus dominios con el software Pfam que se resaltan en la Figura 4A. IMPα también se ha estudiado en tripanosomátidos y se encuentra anotada en la base de datos TriTrypDB con los números de acceso de T. cruzi (TcCLB.509965.110), T. brucei (Tb927.6.2640) y L. major (LmjF.30.1120) (Canela-Pérez et al., 2020).
Importina β
Importina beta es una proteína receptora soluble que se une a Imporina alfa unida a la proteína cargo con una NLS. Esta proteína pertenece a la familia de las carioferinas y se une mediante su extremo amino al extremo amino de importina alfa. La IMPβ de E. histolytica no ha sido estudiada, sin embargo, se ha podido dilucidar la estructura completa en modelo murino (Cingolani et al., 1999). IMPβ está constituida por dominios HEAT consecutivos que conforman una super hélice. Hacia su extremo carboxilo tiene el sitio de unión a Ran GTP, que al unirse genera un cambio alostérico en IMPβ para liberar al complejo trimérico de importación IMPα/IMPβ/Cargo en el nucleoplasma. La plasticidad estructural de IMPβ y su sustrato IMPα es una característica general de las carioferinas (Chook and Blobel; 2001). Se ha descrito que algunas proteínas pueden unirse directamente a IMPβ sin la necesidad de IMPα, como es el caso de la proteína relacionada a la hormona paratiroide (PTHrP) uniéndose a su región amino de la misma manera que la ciclina B1 (Lam et al., 1999; Moore et al., 1999). Estos datos sugieren que IMPβ puede tener más de un sustrato y por lo tanto reconocer diversos tipos de NLSs. Asimismo logramos modelar IMPβ de E. histolytica usando el programa alphafold en la que se resaltan los dominios de unión a IMPα y Ran-GTP (Figura 4B).
La vía de importación nuclear de proteínas de las importinas α/β es un terreno inexplorado en E. histolytica.
Hasta la fecha no se ha estudiado la via α/β de importación de proteínas nucleares en E. histolytica, por lo que nuestra búsqueda de los genes codificantes para IMPα e IMPβ en la base de datos AmoebaDB, abre las puertas a explorar esta vía hasta ahora sin estudiar en este parásito protozoario. Nuestros modelos estructurales indican una alta conservación estructural con las proteínas ortólogas de S. cerevisiae y tripanosomátidos, en donde observamos que se mantienen presentes los dominios principales. Los números de acceso para EhIMPα es EHI_025350 y para EhIMPβ EHI_036520, respectivamente. Nuestros datos indican que esta vía de importación nuclear se encuentra conservada en E. histolytica y en otros parásitos protozoarios como Trypanosoma cruzi y Trypanosoma brucei en donde se han logrado estudiar e identificar estos componentes (Canela-Pérez et al., 2019; Canela-Pérez et al., 2020). La conservación de los factores de esta vía de importación nuclear en eucariontes unicelulares, nos indica un surgimiento temprano en el proceso de importación nuclear de proteínas en una etapa inicial de la evolución de los eucariontes.
Figura 4. A) Estructura de EhIMPα en donde se muestran en color azul cielo los 9 dominios armadillo que sirven de reconocimiento para la estructura de las NLSs; en negro se aprecia el dominio de unión a IMPβ hacia el extremo amino de EhIMPα. B) Estructura de EhIMPβ. En azul señala el sitio de unión a la proteína Ran-GTP, mientras que en púrpura se indica el sitio de unión a IMPα. En cada proteína se indica el extremo amino terminal (NT) y carboxilo teminal (CT).
Conclusión
La vía de transporte nuclear clásica en E. histolytica es un campo inexplorado, por lo que la identificación de las importinas α y β resultan una contribución muy importante al campo del conocimiento científico en este protozoario. La estructura molecular de ambas importinas nos indica una conservación estructural al ser comparada con otros organismos como levaduras, trypanosomas y plantas, resultando necesaria su cristalización para poder ahondar en estudios de comparación evolutiva y filogenia. Será crucial la investigación de fármacos que puedan bloquear de manera específica el mecanismo de importación y exportación nuclear de proteínas en E. histolytica.
Referencias
Conti E, Uy M, Leighton L, Blobel G, Kuriyan J. (1998), Crystallographic analysis of the recognition of a nuclear localization signal by the nuclear import factor karyopherin alpha. Cell. 94(2):193-204. DOI:10.1016/s0092-8674(00)81419-1.
Fontes MR, Teh T, Kobe B. (2000), Structural basis of recognition of monopartite and bipartite nuclear localization sequences by mammalian importin-alpha. J Mol Biol.297(5):1183-94. DOI:10.1006/jmbi.2000.3642.
Avalos-Padilla, Betanzos A., Javier-Reyna R., García-Rivera G., Chávez-Munguía B., Lagunes-Guillén A., Ortega J., Orozco E. (2015). EhVps32 Is a Vacuole-Associated Protein Involved in Pinocytosis and Phagocytosis of Entamoeaba histolytica. PLoS Pathog. 11(7):e1005079. doi: 10.1371/journal.ppat.1005079.
Becker SM, Cho KN,* Xiaoti Guo X,* Kirsten Fendig K,† Oosman MN, Whitehead R, Cohn SM, Houpt (2010). Epithelial Cell Apoptosis Facilitates Entamoeba histolytica Infection in the Gut. Am J Pathol. 176(3): 1316–1322. doi: 10.2353/ajpath.2010.090740
Begum S, Gorman H, Chadha A, Chadee K. (2021). Entamoeba histolytica. Trends Parasitol. 37(7):676-677. doi: 10.1016/j.pt.2021.01.001.
Begum, Quach J., Chadee K. (2015). Immune Evasion Mechanisms of Entamoeba histolytica: Progression to Disease. Front Microbiol. 6: 1394. doi: 10.3389/fmicb.2015.01394.
Bernardes NE, Takeda AA, Dreyer TR, Freitas FZ, Bertolini MC, Fontes MR. (2015). Structure of Importin-α from a Filamentous Fungus in Complex with a Classical Nuclear Localization Signal. PLoS One.10(6):e0128687. DOI:10.1371/journal.pone.0128687.
Berninghausen, Leippe M., (1997). Necrosis versus apoptosis as the mechanism of target cell death induced by Entamoeba histolytica. Infect Immun. 65(9):3615-21.doi: 10.1128/iai.65.9.3615-3621.1997.
Bischoff FR, Klebe C, Kretschmer J, Wittinghofer A, Ponstingl H. (1994), RanGAP1 induces GTPase activity of nuclear Ras-related Ran. Proc Natl Acad Sci U S A. 91(7):2587-91. DOI:10.1073/pnas.91.7.2587.
Bischoff FR, Ponstingl H. (1991), Catalysis of guanine nucleotide exchange on Ran by the mitotic regulator RCC1. Nature. 354(6348):80-2. DOI:10.1038/354080a0.
Canela-Pérez I, López-Villaseñor I, Cevallos AM, Hernández R. (2018). Nuclear distribution of the Trypanosoma cruzi RNA Pol I subunit RPA31 during growth and metacyclogenesis, and characterization of its nuclear localization signal. Parasitol Res.117(3):911-918. DOI:10.1007/s00436-018-5747-4.
Canela-Perez, I.; Lopez-Villasenor, I.; Cevallos, A.M.; Hernandez, R. Trypanosoma cruzi Importin alpha: ability to bind to a functional classical nuclear localization signal of the bipartite type. Parasitol Res 2020, 119, 3899-3907, doi:10.1007/s00436-020-06885-z.
Casanova M, Portalès P, Blaineau C, Crobu L, Bastien P, Pagès M.(2008), Inhibition of active nuclear transport is an intrinsic trigger of programmed cell death in trypanosomatids. Cell Death Differ.15(12):1910-20. DOI: 10.1038/cdd.2008.132.
Cautain B, Hill R, de Pedro N, Link W. (2015). Components and regulation of nuclear transport processes. FEBS J.282(3):445-62. DOI:10.1111/febs.13163.
Chang CW, Couñago RL, Williams SJ, Bodén M, Kobe B. (2012). Crystal structure of rice importin-α and structural basis of its interaction with plant-specific nuclear localization signals. Plant Cell. 24(12):5074-88. DOI: 10.1105/tpc.112.104422.
Chang CW, Williams SJ, Couñago RM, Kobe B.(2014). Structural basis of interaction of bipartite nuclear localization signal from Agrobacterium VirD2 with rice importin-α. Mol Plant. 7(6):1061-1064. DOI: 10.1093/mp/ssu014.
Christy NCV, Petri Jr (2011). Mechanisms of adherence, cytotoxicity and phagocytosis modulate the pathogenesis of Entamoeba histolytica. Future Microbiol. 6(12):1501-19. doi: 10.2217/fmb.11.120.
Cingolani, Petosa C., Weis K., Müller CW. (1999). Structure of importin-beta bound to the IBB domain of importin-alpha. Nature. 399(6733):221-9. doi: 10.1038/20367.
Conti E, Kuriyan J. (2000). Crystallographic analysis of the specific yet versatile recognition of distinct nuclear localization signals by karyopherin alpha. Structure. 15;8(3):329-38. DOI:10.1016/s0969-2126(00)00107-6.
Debnath, Parsonage D., Andrade RM, He C., Cobo ER, Hirata K., Chen S., García-Rivera G., Esther Orozco, Martínez MB., Gunatilleke SS., Barrios AM, Arkin MR, Leslie B Poole LB., McKerrow JH, Reed SL. (2012). A high-throughput drug screen for Entamoeba histolytica identifies a new lead and target. Nat Med. 18(6):956-60. doi: 10.1038/nm.2758.
Dias SM, Wilson KF, Rojas KS, Ambrosio AL, Cerione RA. (2009). The molecular basis for the regulation of the cap-binding complex by the importins. Nat Struct Mol Biol.16(9):930-7. DOI: 10.1038/nsmb.1649.
Fontes MR, Teh T, Jans D, Brinkworth RI, Kobe B. (2003). Structural basis for the specificity of bipartite nuclear localization sequence binding by importin-alpha. J Biol Chem. 278(30):27981-7. DOI:10.1074/jbc.M303275200.
Fournier D, Palidwor GA, Shcherbinin S, Szengel A, Schaefer MH, Perez-Iratxeta C, Andrade-Navarro MA. (2013), Functional and genomic analyses of alpha-solenoid proteins. PLoS One. 8(11):e79894. DOI:10.1371/journal.pone.0079894.
Goldfarb D., Corbett A., Mason D., Harreman M., Adam S. (2004). Importin a: a multipurpose nuclear-transport receptor. Trends Cell Biol.14 (9): 505-514. DOI:10.1016/j.tcb.2004.07.016.
Görlich D, Panté N, Kutay U, Aebi U, Bischoff FR. (1996). Identification of different roles for RanGDP and RanGTP in nuclear protein import. EMBO J. 15(20):5584-94. DOI: https//doi.org/10.1002/j.1460-2075.1996.tb00943.x.
Hanadate, Saito‐Nakano Y., Nakada Tsukui K, Nozaki T. (2016). Endoplasmic reticulum‐resident Rab8A GTPase is involved in phagocytosis in the protozoan parasite Entamoeba histolytica. Cell Microbiol.18(10): 1358–1373. doi: 10.1111/cmi.12570
Haque, Huston CD, Hughes M, Houpt E, Petri Jr WA. (2003). Amebiasis. N Engl J Med. 348(16):1565-73. doi: 10.1056/NEJMra022710.
Hoelz A, Blobel G. (2004). Cell biology: popping out of the nucleus. Nature.432(7019):815-6. DOI:10.1038/432815a
Jovanovic-Talisman T, Tetenbaum-Novatt J, McKenney AS, Zilman A, Peters R, Rout MP, Chait BT. (2009), Artificial nanopores that mimic the transport selectivity of the nuclear pore complex. Nature.457(7232):1023-7. DOI: 10.1038/nature07600.
Kimura, Nakamura T., Nawa Y. (2007), Experience with intravenous metronidazole to treat moderate-to-severe amebiasis in Japan, Am J Trop Med Hyg., 77(2):381-5.
King MC, Lusk CP, Blobel G. (2006). Karyopherin-mediated import of integral inner nuclear membrane proteins. 442(7106):1003-7. DOI:10.1038/nature05075.
Kobe B.(1999). Autoinhibition by an internal nuclear localization signal revealed by the crystal structure of mammalian importin alpha. Nat Struct Biol. 6(4):388-97. DOI:10.1038/7625.
Kutay U, Bischoff FR, Kostka S, Kraft R, Görlich D. (1997). Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell.90(6):1061-71. DOI:10.1016/s0092-8674(00)80372-4.
Lam MH, Briggs LJ, Hu W, Martin TJ, Gillespie MT, Jans (1999). Importin beta recognizes parathyroid hormone-related protein with high affinity and mediates its nuclear import in the absence of importin alpha. J Biol Chem. 274(11):7391-8.doi: 10.1074/jbc.274.11.7391.
Leung SW, Harreman MT, Hodel MR, Hodel AE, Corbett AH. (2003). Dissection of the karyopherin alpha nuclear localization signal (NLS)-binding groove: functional requirements for NLS binding. J Biol Chem. 278(43):41947-53. DOI:10.1074/jbc.M307162200.
Lopez-Reyez I., Garcia-Rivera G., Banuelos C., Herranz S., Vincent O., Lopez-Camarillo C., Marchat LA., Orozco E. (2010). Detection of the endosomal sorting complex required for transport in Entamoeba histolytica and characterization of the EhVps4 protein. Journal of biomedicine and biotechnology. 890674
Ma J, Goryaynov A, Yang W. (2016). Super-resolution 3D tomography of interactions and competition in the nuclear pore complex. Nat Struct Mol Biol. 23(3):239-47. DOI: 10.1038/nsmb.3174.
Marie C and Petri Jr. WA (2013). Amoebic dysentery. BMJ Clin Evid. 2013:0918. PMID: 23991750
Matunis MJ, Coutavas E, Blobel G. (1996). A novel ubiquitin-like modification modulates the partitioning of the Ran-GTPase-activating protein RanGAP1 between the cytosol and the nuclear pore complex. J Cell Biol.135(6 Pt 1):1457-70. DOI:10.1083/jcb.135.6.1457.
Moore, Yang J., Truant R., Kornbluth S. (1999). Nuclear Import of Cdk/Cyclin Complexes: Identification of Distinct Mechanisms for Import of Cdk2/Cyclin E and Cdc2/Cyclin B1. J Cell Biol.144(2): 213–224. doi: 10.1083/jcb.144.2.213
Moroianu J, Blobel G, Radu A. (1996). Nuclear protein import: Ran-GTP dissociates the karyopherin alphabeta heterodimer by displacing alpha from an overlapping binding site on beta. Proc Natl Acad Sci U S A. 93(14):7059-62. DOI:10.1073/pnas.93.14.7059.
Oka M, Yoneda Y. (2018). Importin α: functions as a nuclear transport factor and beyond. Proc Jpn Acad Ser B Phys Biol Sci.94(7):259-274. DOI:10.2183/pjab.94.018.
Pritt, Clark CG., (2008), Amebiasis, Mayo Clin Proc., 83(10):1154-9; quiz 1159-60.doi: 10.4065/83.10.1154.
Ralston (2015).Chew on this: Amoebic trogocytosis and host cell killing by Entamoeba histolytica. 31(9): 442–452. doi: 10.1016/j.pt.2015.05.003
Ralston KS., Solga MD, Mackey-Lawrence NM, Somlata Bhattacharya A., Petri Jr., WA (2014). Trogocytosis by Entamoeba histolytica contributes to cell killing and tissue invasion. Nature. 508 (7497), 526-530. doi: 10.1038/nature13242.
Riggleman B, Wieschaus E, Schedl P.(1989). Molecular analysis of the armadillo locus: uniformly distributed transcripts and a protein with novel internal repeats are associated with a Drosophila segment polarity gene. Genes Dev.3(1):96-113. DOI:10.1101/gad.3.1.96.
Ritterhoff T, Das H, Hofhaus G, Schröder RR, Flotho A, Melchior F. (2016). The RanBP2/RanGAP1*SUMO1/Ubc9 SUMO E3 ligase is a disassembly machine for Crm1-dependent nuclear export complexes. Nat Commun.7:11482. DOI: 10.1038/ncomms11482.
Sahoo N., Labruyere E., Bhattacharya S., Sen P., Guillen N., Battacharya A. (2004). Calcium binding protein 1 of the protozoan parasite Entamoeba histolytica interacts with actin and is involved in cytoskeleton dynamics. Journal of Cell Science 117(Pt 16), 3625-3634.
Saito-Nakano Y, Loftus BJ, Hall N, Nozaki (2005). The diversity of Rab GTPases in Entamoeba histolytica. Exp Parasitol. 110(3):244-52. doi: 10.1016/j.exppara.2005.02.021.
Seewald MJ, Körner C, Wittinghofer A, Vetter IR. (2002). RanGAP mediates GTP hydrolysis without an arginine finger. Nature.415(6872):662-6. DOI:10.1038/415662a.
Sha SM., Usmanghani K., Akhtar N., Akram M., Asif HM., Hasan MM., (2016). Clinical study on the efficacy of amoebex (coded herbal drug) compared with metronidazole for the treatment of amoebic dysentery. Pakistan Journal of Pharmaceutical Sciences 29(6), 2005-2014.
Singh, Walia AK, Kanwar JR., (2016). Protozoa lectins and their role in host-pathogen interactions. 34(5):1018-1029.doi: 10.1016/j.biotechadv.2016.06.002.
Somlata, Bhattacharya, Bhattacharya A., (2011)., A C2 domain protein kinase initiates phagocytosis in the protozoan parasite Entamoeba histolytica. Nat Commun. 2:230. doi: 10.1038/ncomms1199.
Somlata, Kamanna, Agrahari M., Babuta M., Bhattacharya S., Bhattacharya A. (2012). Autophosphorylation of Ser428 of EhC2PK plays a critical role in regulating erythrophagocytosis in the parasite Entamoeba histolytica. J Biol Chem. 287(14):10844-52. doi: 10.1074/jbc.M111.308874.
Stanley Jr, (2003), Amoebiasis, Lancet, 361(9362):1025-34. doi: 10.1016/S0140-6736(03)12830-9.
Stewart M, Rhodes D. (1999). Switching affinities in nuclear trafficking. Nat Struct Biol.6(4):301-4. DOI:10.1038/7529.
Stewart M. (2000). Insights into the molecular mechanism of nuclear trafficking using nuclear transport factor 2 (NTF2). Cell Struct Funct. 25(4):217-25. DOI:10.1247/csf.25.217.
Stewart M. (2007). Molecular mechanism of the nuclear protein import cycle. Nat Rev Mol Cell Biol.8(3):195-208. DOI:10.1038/nrm2114
Strambio-De-Castillia C, Niepel M, Rout MP. (2010). The nuclear pore complex: bridging nuclear transport and gene regulation. Nat Rev Mol Cell Biol.11(7):490-501. DOI: 10.1038/nrm2928.
Verma, Nozaki T., Datta S. (2016). Role of EhRab7A in phagocytosis of type 1 fimbriated E. coli by Entamoeba histolytica. Mol Microbiol. 102(6):1043-1061. doi: 10.1111/mmi.13533.
Verma, Saito-Nakano Y., Nozaki T., Datta S. (2015). Insights into endosomal maturation of human holo-transferrin in the enteric parasite Entamoeba histolytica: essential roles of Rab7A and Rab5 in biogenesis of giant early endocytic vacuoles. Cellular microbiology. 17(12), 1779-1796. doi.org/10.1111/cmi.12470
Vetter IR, Nowak C, Nishimoto T, Kuhlmann J, Wittinghofer A. (1999). Structure of a Ran-binding domain complexed with Ran bound to a GTP analogue: implications for nuclear transport. 398(6722):39-46. DOI:10.1038/17969.
Wagstaff KM, Sivakumaran H, Heaton SM, Harrich D, Jans DA. (2012). Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV and dengue virus. Biochem J. 443(3):851-6. DOI: 10.1042/BJ20120150.
Wirthmueller L, Roth C, Fabro G, Caillaud MC, Rallapalli G, Asai S, Sklenar J, Jones AM, Wiermer M, Jones JD, Banfield MJ. (2015). Probing formation of cargo/importin-α transport complexes in plant cells using a pathogen effector. Plant J.81(1):40-52. DOI: 10.1111/tpj.12691.
Xu L, Massagué J. (2004), Nucleocytoplasmic shuttling of signal transducers. Nat Rev Mol Cell Biol. 5(3):209-19. DOI:10.1038/nrm1331.
Yang SN, Takeda AA, Fontes MR, Harris JM, Jans DA, Kobe B. (2010). Probing the specificity of binding to the major nuclear localization sequence-binding site of importin-alpha using oriented peptide library screening. J Biol Chem. 285(26):19935-46. DOI: 10.1074/jbc.M109.079574.
Más de una vez, a lo largo de nuestra vida, nos ha picado una abeja y de inmediato sentimos dolor. ¿Es por el aguijón que se nos quedó enterrado? La verdad no. Tal dolor es una respuesta de nuestro sistema nervioso por un exceso de inflamación en las células de la piel, producido por el veneno que la abeja nos inyectó. Éste y otros venenos se componen, entre otras cosas, de una enzima de apenas 14-16 kDa (kilodaltons) que se llama Fosfolipasa A2, o PLA2 por sus siglas en inglés. Esta pequeña enzima, o proteína, funge como el agente activo que desencadena el proceso inflamatorio y corresponde a cerca del 15% del peso seco del veneno.
La PLA2 tiene la habilidad particular de cortar fosfolípidos. Específicamente, corta la cola derecha (sn-2) de éstos (Fig. 1). Los fosfolípidos, dicho sea de paso, son moléculas con un extremo polar o cabeza (que le gusta el agua) y una cola no-polar (o hidrófoba, que evita el agua). Son los responsables de generar una barrera llamada “membrana”, que da soporte a receptores proteicos presentes en todo organelo, toda célula, y en todo tejido de nuestro cuerpo, por ejemplo, en las células de la piel. Así que la abeja, asustada por el instinto de una amenaza, nos inyecta un “shot” de enzima y ésta, al instante, comienza a realizar la función para la que fue diseñada: cortar colas derechas de fosfolípidos desencadenando un mecanismo inflamatorio en las células circunvecinas. Un verdadero pandemónium molecular. Cabe señalar que todos los venenos, de serpiente, araña, abeja, avispa, entre otros, cuentan con este poderoso corta-lípidos, donde la especie busca paralizar a su presa por un exceso de inflamación, o bien, usarlo como mecanismo de defensa cuando se ve amenazada.
Figura 1. La Fosfolipasa A2 de tipo secretada (sPLA2) corta la cola sn-2 de un fosfolípido de la membrana plasmática, la cual corresponde al ácido graso poliinsaturado: ácido araquidónico. Este ácido graso puede ser procesado posteriormente hasta por 3 tipos de enzimas, donde particularmente la ciclooxigenasa (COX1 o COX2) se encarga de transformarlo en prostaglandinas, las cuales participan como mediadoras del proceso fisiológico de inflamación, dolor y fiebre.
Puesto que evolutivamente la PLA2 se ha convertido en una máquina perpetua y eficiente de corte, ésta corta sin parar la cola derecha de tantos fosfolípidos como le sea posible en las células cercanas donde fue inyectada. Después del dolor viene una hinchazón, producto de la inflamación, donde un colectivo de agentes del sistema inmune se reúne para revisar la zona y tratar de detener el festín de la PLA2. La alta sensibilidad de una región del cuerpo inflamada, tal que el mínimo tacto en la zona genera un gran dolor, es algo que hemos sufrido.
Por alguna extraña razón, interesante y aún bajo estudio, en la fisiología humana necesitamos de un “barbero” de lípidos, donde no solo contamos con la PLA2 sino que también producimos otras primas hermanas de ésta (e.g. PLA1, PLC y PLD), con la encomienda de cortar distintas zonas del fosfolípido, donde a su vez, cada producto regula determinado proceso celular de forma única. Y, por si fuera poco, en nuestro cuerpo contamos con diversos tipos de PLA2, cuyas diferencias estructurales tan sutiles les permiten cortar colas de lípidos de distintos tipos y en variados contextos celulares y tejidos de nuestro cuerpo. Gracias a estos mecanismos, podríamos afirmar que tenemos micro-periodos de inflamación y desinflamación donde la PLA2 actúa de una forma sumamente regulada. Cuando por factores externos ocurre un desbalance en la producción de PLA2, caemos en una inflamación constante o crónica. Entonces se activan las alarmas. Una inflamación crónica en el sistema nervioso o cerebro puede presentarse en las enfermedades de Alzheimer, migraña o epilepsia. La inflamación en el tejido muscular se presenta en la esclerosis múltiple, mientras que en la piel (el órgano más grande del cuerpo) da lugar a dermatitis atópica, psoriasis, calvicie, entre otras. Hasta una hernia discal está asociada con la PLA2. En pocas palabras, un exceso de inflamación no siempre se asocia a la picadura de una abeja.
La hidrólisis de fosfolípidos producida por la enzima PLA2 es solo el primer paso de una ruta metabólica que termina en la síntesis de ácido araquidónico (AA), formado por una cadena de 20 átomos de carbono y 5 dobles enlaces entre éstos (Fig. 1). Este ácido graso poliinsaturado o PUFA (por sus siglas en inglés), genera moléculas proinflamatorias al ser metabolizado. Por lo tanto, es importante aclarar que una actividad de la PLA2 no siempre implica inflamación, pues se requiere una acumulación de AA para desencadenar el proceso.
En lo que respecta a los venenos, no todo es dañino. Se sabe que la PLA2 de abeja, así como de algunas otras especies, en la dosis adecuada, puede tener efectos irónicamente terapéuticos de tipo antimicrobiano, antitumoral, anti-inflamatorio, anti-neurodegenerativo, antinociceptivo, entre otros. El truco está en que si la PLA2 de abeja gusta de cierto tipo de fosfolípidos presentes en una bacteria (e.g. E.Coli), entonces probabilísticamente esta bacteria sería cortada en mayor proporción que nuestras células. Dicho de otra forma, una micro-inflamación pudiera generar estragos a determinados microrganismos invasores, células de cáncer, o poner orden en determinadas regiones de una célula propia del cuerpo humano.
Por otra parte, ¿qué decir de aquellos compuestos que pueden regular la función de dichas PLA2? Evolutivamente, las especies han generado agentes que acompañan a la PLA2 en los venenos. Sin embargo, aún falta mucho por comprender el rol que juega cada uno y la sinergia que presentan al momento de inyectarse juntos. En el caso de la abeja, se sabe que una pequeña molécula llamada “melitina” permite potenciar la acción de corte (inflamatoria) de la PLA2. Del mismo modo, se sospecha que otros péptidos presentes en el veneno, que generan poros en la membrana, tienen una acción biológica conjunta para atacar por todos lados a una presa, o defenderse de algún depredador.
Las plantas también han desarrollado evolutivamente algunos biocompuestos (como los polifenoles) para defenderse de algunas plagas de microrganismos, mientras que otros depredadores serán atraídos para consumir el fruto, y luego de una buena digestión, ayudar a la planta a esparcir su semilla en otros lugares. Así que, cuando el ser humano consume cierto tipo de plantas (raíz, semilla, tallo, fruto, flor, hoja o resina), algunos biocompuestos pueden presentar un efecto anti-inflamatorio sorpredente.
De lo anterior, se desprende que aquellos biocompuestos con propiedades anti-inflamatorias como el resveratrol, el CBD, la quercetina, el ácido oleanólico, tienen la propiedad de inhibir a la enzima que desencadena el proceso inflamatorio en la membrana celular. Recientemente, dimos un pequeño gran paso en la comprensión de la actividad de la PLA2 y la regulación de una variedad de biocompuestos naturales y sintéticos con propiedades anti-inflamatorias. Encontramos que, contra lo esperado, no todos los anti-inflamatorios son capaces de inhibir el efecto de corte. Algunos de éstos, al contrario, lo aceleran. Esto pudiera indicar, por ejemplo, que en un sistema celular deberían de sobre-inflamar. Y más bien esto sería la causa del buen resultado que muestran frente a células cancerígenas, donde ya per se equivale a un ambiente inflamatorio. Además, en este trabajo experimental y computacional, proponemos un posible mecanismo de acción donde biocompuestos de naturaleza química hidrófoba pueden modular la acción de esta enzima a través la regulación de las propiedades físico-químicas de la membrana celular.
Una ingeniería de fármacos que use a su favor la gran riqueza de la naturaleza para modular, “al gusto”, la acción de los venenos y generar estrategias terapéuticas para contrarrestar la inflamación crónica en el cuerpo humano, pudiera desprenderse como consecuencia del estudio biofísico y preliminar que recién hemos publicado en Langmuir 2024, 40, 13, 7038–7048 (https://pubs.acs.org/doi/abs/10.1021/acs.langmuir.4c00084).
Foto de portada: Pexels por David Hablützel
Árboles: alimento y algo más en la producción de ovinos a pequeña escala en Tabasco
La cría de ovinos o borregos es una actividad con gran presencia en las zonas rurales de México. En el país se tienen registradas aproximadamente 420 mil unidades de producción de esta especie, entre las que predominan las de tipo familiar orientadas principalmente a obtener carne, producto ampliamente demandado y cuyo precio promedio de venta en 2023 fue de $35.99/kg para ganado en pie, y de $92.16/kg para ganado en canal. A pesar de la consolidada demanda y del buen precio de venta del cordero, la producción es insuficiente para abastecer el consumo nacional, siendo necesario importar aproximadamente el 60% de la carne ovina que se requiere.
No obstante la importancia social y económica de este sector, durante los últimos diez años la producción nacional de carne de ovino ha crecido apenas un 1.25%, con un inventario de 8,805,206 cabezas a lo largo del territorio nacional; y cuya contribución del estado de Tabasco es del 1%, con un crecimiento promedio anual del 0.17% (SIAP, 2023). El bajo crecimiento en el inventario estatal se debe a múltiples factores, entre los que se pueden mencionar: un bajo uso de tecnología en las explotaciones de tipo familiar, ausencia de organización de los productores, inseguridad en los sitios de producción, y presencia de enfermedades que disminuyen la eficiencia productiva o que comprometen la vida o el estado de salud de los animales.
Sin embargo, la superficie con uso y vocación para agostadero, pastos naturales, arbustos, hierbas, matorrales, o enmontada, que no tiene uso agrícola, pero que puede ser aprovechada para la alimentación del ganado, representa el 40% de la superficie total del estado. Esto puede favorecer la cría de ovinos, actividad que se desarrolla principalmente en sistemas de producción conocidos como extensivos, los cuales se identifican porque tratan de aprovechar al máximo la pradera mediante libre pastoreo (Figura 1). En este sistema, el pasto es la principal fuente de nutrientes (proteína y energía) para los ovinos en sus diferentes etapas productivas. Sin embargo, la disponibilidad de pasto en la pradera es variable en cantidad y en contenido de nutrientes a lo largo del año, lo que propicia que no se cubran las necesidades alimenticias de los animales y se vea limitado su potencial productivo (Nuncio-Ochoa et al., 2001).
Además, para la cría de ovinos en una unidad de producción de tipo familiar es común el uso de animales cruzados, en los cuales la presencia de pelo y lana en el cuerpo es una característica frecuente; los rebaños son pequeños (menos de 30 ejemplares), se invierte poco capital en el sistema de producción y se dedican menos horas para el manejo y cuidado de los animales. En las áreas de pastoreo generalmente la cubierta vegetal consiste en algún tipo de pasto o mezcla de diferentes pastos, y aunque pueden existir diversas clases de árboles, el uso de su follaje, aun cuando es apetecible para los ovinos, no es una práctica común en la alimentación de los animales. Esto se puede atribuir, por un lado, a que el productor desconoce el valor nutritivo del follaje, y por otro, a la dificultad para que los ovinos puedan acceder a las hojas de los árboles, así como a la palatabilidad que cada uno tiene, es decir, qué tan agradable es como para que pueda ser consumido.
Considerando que en el estado de Tabasco existe potencial para el aprovechamiento de diferentes especies de árboles que mejoren el sistema de producción ovina, es de interés el generar información que contribuya a ampliar el conocimiento de los productores sobre los beneficios que pueden proporcionar los árboles para dar sostenibilidad a la cría ovina. Por esta razón, un grupo de investigación del Instituto Nacional de Investigaciones, Forestales, Agrícolas y Pecuarias-Campo Experimental Huimanguillo, está conduciendo estudios sobre el uso de follaje de árboles como complemento alimenticio de los ovinos, con énfasis en cocoíte y moté, por ser leguminosas arbóreas con alta presencia en la región tropical húmeda; y simultáneamente, está identificando la viabilidad técnica y económica de diversos componentes tecnológicos en un sistema de producción silvopastoril a pequeña escala.
Como parte de la información que se ha generado al respecto en dichos estudios, en este escrito se abordan las ventajas y desventajas de incorporar árboles proveedores de follaje en las unidades de producción de ovinos de tipo familiar, así como los aspectos básicos que se deben considerar para el manejo intensivo de estos árboles en la modalidad de cosecha manual del follaje, acarreo y ofrecimiento en verde en pesebre (Figura 2), la cual está dentro de los sistemas silvopastoriles. También se indican algunos ejemplos de árboles con posibilidades de manejo bajo este sistema y se enfatiza el beneficio de los árboles como proveedores de protección para dar más confort a los animales en pastoreo.
Figura 1. Unidades de producción de ovinos de tipo familiar en Huimanguillo, Tabasco. En este tipo de unidades de producción el principal alimento de los ovinos lo representa el pasto, y el uso de follaje de árboles como alimento es mínimo. Fotografías por Jorge Oliva Hernández.
Figura 2. Árboles de cocoíte con manejo intensivo para obtener follaje (izquierda); cosecha manual de follaje de cocoíte (centro); ovejas consumiendo follaje de cocoíte (derecha). Fotografías por Jorge Oliva Hernández
Ventajas del uso de árboles forrajeros en la unidad familiar de producción ovina
Dar apoyo y sostenibilidad a la producción en pastoreo
La ganadería extensiva en la región tropical se ha relacionado con daño a sus zonas boscosas, debido a que éstas se han sustituido por praderas para el pastoreo de ganado, con el consecuente deterioro de suelo y la pérdida de la biodiversidad asociada a estos ecosistemas. Por tal motivo, en décadas recientes, se ha procurado promover acciones que transformen los sistemas de producción ganaderos de tipo extensivo hacia sistemas silvopastoriles, donde además de estar presente el ganado, también están presentes árboles que proporcionan diversos productos y servicios ecosistémicos (forraje, frutos, madera, cercos, sombra, protección, entre otros), arbustos forrajeros y pastos principalmente. En los sistemas extensivos, comúnmente el productor de tipo familiar no tiene divisiones de las áreas donde pastorea el ganado, lo que dificulta el control de los días que los animales permanecerán pastando y los días de descanso de la pradera para permitir el rebrote del pasto y que éste pueda seguir disponible en cantidad y calidad durante todo el año. Además, el consumo voluntario de pasto no es suficiente para cubrir las necesidades de nutrientes que tienen los animales, situación que justifica el uso de follaje de árboles como complemento alimenticio.
En el sistema de producción silvopastoril con el ganado, están presentes tres grandes componentes principales: árboles, pradera y animales, los cuales interaccionan entre sí, y cualquier desequilibrio en el manejo de uno de los componentes, afecta a los otros dos. Así, el productor tendrá que definir cuál de los tres componentes tendrá prioridad sobre los otros, procurando que su aprovechamiento no deje fuera a ninguno de los componentes y se rompa el equilibrio del sistema. En el sistema silvopastoril con un manejo intensivo de los árboles, se deben incluir podas y la cosecha manual del follaje, por lo cual, resulta importante conocer los tipos de árboles cuyo follaje puede ser consumido con facilidad por los ovinos y cosechado de forma manual fácil y segura.
Complementar la dieta de ovinos en pastoreo
Durante el pastoreo, los ovinos están expuestos a diversas condiciones, como temperaturas elevadas, lluvia, viento y también parásitos, lo que hace necesario que parte de los nutrientes que obtienen mediante el consumo de pasto, los destinen a funciones que les permitan sobrevivir y mantener un equilibrio, reduciendo con ello la cantidad de nutrientes requeridos para funciones productivas como el crecimiento del animal. Por lo anterior, es inevitable ofrecer un complemento alimenticio, que puede ser el follaje de algunos árboles, porque representa una fuente de proteína y fibra que puede ser digerida y aprovechada.
Árboles que se pueden incorporar a la unidad de producción ovina
Dentro de los árboles más estudiados debido a la composición química de su follaje y su fácil consumo por lo ovinos, están las leguminosas como el cocoíte y el moté. Además, existen otros árboles que no son leguminosas, que pueden utilizarse como complemento alimenticio (Cuadro 1, Figura 3).
Cuadro 1. Ejemplos de árboles cuyo follaje se puede utilizar como alimento para ovinos
Figura 3. Hojas de algunos árboles en fincas ganaderas localizadas en la región tropical húmeda y cuyo follaje es consumido con facilidad por los ovinos. A. Moté (Erythrina americana Mill.); B. Cocoíte (Gliricidia sepium (Jacq.) Walp.); C. Morera (Morus alba L); D. Tulipán africano (Spathodea campanulata).
Manejo intensivo de árboles proveedores de follaje
La incorporación de árboles proveedores de follaje en la unidad de producción familiar, se facilita si forman parte del cerco vivo y se someten a un manejo intensivo, que consiste en sembrarlos a una distancia de un metro entre árboles; realizar podas controladas a intervalos de entre 80 y 100 días; y no permitir que el árbol tenga una altura superior a 1.8 metros, con el fin de facilitar la cosecha manual del follaje. Los árboles que tienen mayor posibilidad de incorporarse al sistema de producción silvopastoril, son aquellos cuya siembra se puede realizar por material vegetal usando estacas o varetas, puesto que la producción de follaje empezará más rápido en comparación con árboles sembrados mediante semilla. Árboles de moté, cocoíte y morera, son ejemplos de especies que se pueden sembrar usando varetas. En el caso de los árboles de guásimo y tulipán africano, solo se pueden sembrar por semilla.
La cosecha manual de follaje se ha estudiado en cocoíte y moté, y en promedio un productor puede cosechar entre ocho y nueve kilogramos de follaje verde en una hora de trabajo. No obstante, se debe considerar que la eficiencia de cosecha es influida por el diámetro del árbol, época del año y tipo de árbol (Castillo et al., 2021). En los árboles proveedores de follaje no se ha estudiado el uso del estiércol de los ovinos como abono orgánico y su posible influencia sobre el incremento productivo de follaje. Sin embargo, es factible que la respuesta sea positiva, lo que facilitaría el reciclaje del estiércol ovino y la incorporación de árboles al sistema de producción, dando con ello sostenibilidad a la unidad de producción.
El número de árboles que se requieren en una unidad de producción se determina por el número de ovinos en el rebaño y por el nivel de complementación alimenticia que se desee utilizar. Pero es factible que el follaje de los árboles pueda representar entre un 10 y un 30 por ciento del total del alimento que consume un ovino.
Desventajas de la incorporación de árboles forrajeros a la unidad de producción ovina
En cuanto al manejo intensivo de árboles proveedores de follaje en la unidad de producción, es necesario que se consideren las siguientes limitantes: a) se requiere dedicar tiempo y trabajo para el cuidado de los árboles con el fin de que se mantengan libres de malezas y plantas trepadoras porque dificultan el proceso de cosecha; b) una alta densidad de árboles favorece que se resguarden en ellos avispas y orugas, las cuales, en caso de no tomarse medidas preventivas, pueden representar un peligro para la persona encargada de la cosecha de follaje; c) la cosecha de follaje implica un trabajo diario cuando se quiere ofrecer en verde, aunque también existe la posibilidad de darle un tratamiento posterior para usarlo como heno (Castillo et al., 2021 y 2023).
Por último, resaltar que a pesar de las limitantes mencionadas, los árboles proveedores de follaje, además de ser un recurso alimenticio disponible todo el año en la unidad de producción, también aportan otros beneficios: el más relevante para los ovinos es la protección y confort que brinda el área bajo la copa, proveyendo sombra indispensable en días con altas temperaturas durante el pastoreo, o servir para el resguardo durante la lluvia, por lo que representan una oportunidad para dar sostenibilidad al sistema de producción. Además, los árboles proporcionan otros beneficios medioambientales, los cuales, en lo inmediato, no constituyen un beneficio económico para el productor, pero su presencia permite capturar el dióxido de carbono (CO2) de la atmósfera y retenerlo en su estructura por más tiempo que los pastos, situación que reduce la presencia de CO2 en la atmósfera y contribuye a disminuir el calentamiento de la tierra.
Agradecimientos
El presente escrito forma parte del proyecto “Uso de follaje de Erythrina americana y Gliricidia sepium como complemento alimenticio de ovejas reproductoras en pastoreo” con clave SIGI: 1315835078.
Referencias
Castillo L., E.B., López H., M.A., Vélez I., A., y Oliva H., J. (2021). Sistema silvopastoril de cosecha y acarreo como alternativa para la producción ovina sostenible en el trópico húmedo. Revista Mexicana de Ciencias Forestales 12(66): 1-25.
Castillo L., E.B., Vélez I., A., López H., M.A., y Oliva H., J. (2023). Producción ovina silvopastoril. Costo de la mano de obra para el mantenimiento del cerco vivo. Temas de Ciencia y Tecnología 27(79): 51-55.
Nuncio-Ochoa, G., Nahed-Toral, J., Díaz-Hernández, B., Escobedo-Amezcua, F., Salvatierra Izaba, E. B. (2001). Caracterización de los sistemas de producción ovina en el estado de Tabasco. Agrociencia 35(4), 469-477. https://www.redalyc.org/articulo.oa?id=30235411
Perspectivas de la energía solar fotovoltaica y la planeación para la transición energética
Planta Solar Fotovoltaica en Villanueva, Coahuila (Imagen publicada en 2018 por la Secretaría de Energía, México)
La ahora llamada Energía Solar Fotovoltaica inició su desarrollo hace aproximadamente 70 años. El término fotovoltaico se refiere a la producción de un voltaje cuando un dispositivo fotovoltaico es expuesto a la luz. Este tipo de dispositivos electrónicos, conocidos ahora como celdas solares, producen no solo un voltaje, sino también una fotocorriente cuando se colocan bajo una fuente de radiación luminosa, y en particular bajo la radiación solar. Es decir, convierten la energía proveniente de la radiación solar en energía eléctrica (consultar la bibliografía). A los pocos años de haberse iniciado el desarrollo de las celdas solares, fueron la base para proveer energía a los satélites artificiales. Esto sigue siendo un logro que no se ha podido sustituir, por muy diversas razones, aunque ahora los módulos fotovoltaicos utilizados en aplicaciones espaciales se basan en celdas solares mucho más eficientes que en aquellos primeros años.
En sus inicios era impensable utilizar estos dispositivos como fuente de energía eléctrica terrestre, porque su costo era muy alto. Sin embargo, para finales de los años 70 se empezó a prever que se podría convertir en una fuente alterna de energía eléctrica terrestre, y que sus aplicaciones especiales (telecomunicaciones, educación a distancia, albergues infantiles en zonas rurales, casas de campo, etcétera) serían ya costeables. Gradualmente, como consecuencia del desarrollo científico y tecnológico, el costo de la energía fotovoltaica se ha reducido en un factor de por lo menos 400 a 500 veces (figura 1) respecto al inicial, de forma que el costo actual (LCOE) de la energía fotovoltaica es del orden de $0.05 a $0.10 dólares/KWh. Este costo es ya comparable al de la energía eléctrica generada en plantas termo-eléctricas de gas (ciclo combinado). La ventaja de la energía eléctrica fotovoltaica es que su factor de emisión de gases de efecto invernadero es mucho menor que el de las plantas basadas en gas. Esto aminora los problemas relacionados con el efecto de este tipo de gases emitidos a la atmósfera, ayudando así a reducir el posible incremento de temperatura en la tierra, el cual es una preocupación de todos los países en el mundo, ante la creciente demanda de energía eléctrica, como consecuencia del aumento de la actividad humana.
Figura 1. Evolución del costo del Watt pico Fotovoltaico desde 1975 y proyectado hasta 2035 (basado en W. Rickerson, et al. “Residential Prosumers – Drivers and Policy Options”, IEA- Renewable Energy Technology Deployment, June 2014).
Uno de los cuestionamientos que aún tiene la producción de energía eléctrica mediante módulos fotovoltaicos, es que la radiación solar fluctúa a lo largo del día, causando variación (intermitencia) de la potencia eléctrica generada diariamente. Esto desmotiva a los administradores de las redes eléctricas que se utilizan para proveer energía en las ciudades (CFE en México o empresas de “utilities” en otros países), ya que la inyección variable de energía por plantas fotovoltaicas hacia las redes eléctricas, provoca perturbaciones indeseables. La solución a corto plazo (hay otras soluciones posibles a mediano y largo plazo), es que la energía generada fotovoltaicamente no se inyecte directamente a la red sino a partir de baterías, que mantendrían los niveles de voltaje y corriente proporcionados a la red de manera casi constante. Así, la energía fotovoltaica fluctuante se almacenaría en las baterías y éstas despacharían la energía para ser inyectada de forma más estable en la red eléctrica. Esto implica un costo adicional asociado a las baterías, pero nuevamente el avance tecnológico está ayudando a que esto no represente un grave problema, pues las nuevas baterías (basadas en Li y otras en desarrollo) tienen una relación costo/capacidad cada vez menor. Y aún con este costo adicional, la energía eléctrica fotovoltaica con almacenamiento es bajo y comparable al de las plantas basadas en gas, o con otras alternativas energéticas (carbón, nuclear, hidro, etcétera, [figura 2]).
Figura 2. Costo nivelado de la energía eléctrica (LCOE) para diferentes fuentes en 2023 (a partir de datos de Solar Power Europe).
Con base en lo anterior, varios países (en Europa y Asia) han visto que la energía solar fotovoltaica ayudaría a resolver de manera sostenible y simultánea los problemas asociados a la seguridad energética y a la emisión de gases de efecto invernadero. Por eso, en 2022 las nuevas plantas de energía eléctrica instaladas en el mundo, en un 25% fueron plantas de energía fotovoltaica, mientras que las nuevas plantas basadas en energía nuclear o por gas, tuvieron una reducción respecto al año anterior (figura 3). La elección de la energía fotovoltaica en esos países no es fortuita ni una cuestión de moda, sino que se basa en un razonamiento simple: La cantidad de energía promedio que recibimos a partir de la radiación solar sobre la superficie terrestre es aproximadamente de 23,000 TW por año (considerando su aprovechamiento en sólo el 20%), mientras que la demanda mundial de energía actualmente es del orden de 20 TW por año. Es decir, la cantidad de energía solar recibida sobre la superficie terrestre en un año es suficiente para satisfacer 1,000 veces la demanda energética mundial en la actualidad. Más impresionante es el dato de que este recurso energético “renovable” en solo un año produce más energía que la contenida en total en todas las reservas terrestres de energías no-renovables, como gas, petróleo, carbón y uranio (por eso es la fuente para la vida terrestre).
Figura 3. Tasa de crecimiento de nuevas plantas de generación de energía eléctrica durante 2022 (a partir de datos de Solar Power Europe).
En resumen, la energía solar es abundante, ya se puede generar electricidad fotovoltaica de manera económicamente competitiva, reduce la emisión de gases de efecto invernadero y ayuda a resolver los problemas asociados a la inseguridad energética de muchos países. Consecuentemente, solo podemos esperar que en los próximos años la energía solar se convierta en una de las fuentes energéticas con mayor contribución para satisfacer la demanda mundial. Así que lo mejor que se puede hacer en todos los países, pero especialmente en aquellos con economías emergentes y con alto nivel de insolación solar, como México, es iniciar el crecimiento sostenido de la energía solar. Esto requiere de un cambio en las perspectivas de planeación, pues se trata de planear a mediano y largo plazos, más allá de los planes establecidos por administraciones gubernamentales democráticas de corto plazo (planes sexenales). En otras palabras, la planeación energética tiene que ser a mediano plazo, más allá de la duración típica de los regímenes gubernamentales en países democráticos, independientemente de partidos y de los gobernantes en esos periodos.
Debe notarse que aquí no se considera que la energía solar fotovoltaica será la única fuente de energía eléctrica que satisfará la demanda en el futuro. Hay otras alternativas como por ejemplo, la propia energía solar térmica, la geotérmica, la eólica o la hidro-electricidad; lo que se plantea es que la energía solar fotovoltaica, por sus características, proveerá en una gran proporción la energía eléctrica requerida por los diversos países en el futuro. Todas estas alternativas deberán participar, conjuntamente, incluyendo las plantas térmicas de gas a ciclo combinado, durante el periodo de transición energética. Sin embargo, es necesario establecer en qué proporción cada una de estas alternativas energéticas configurarán nuestro portafolio energético en los siguientes años. En cada país este portafolio será diferente, dependiendo de las condiciones específicas en cada uno. Habrá países sin recursos geotérmicos o con bajo nivel de insolación, mientras otros tendrán gran potencial de hidroelectricidad, por lo que la mezcla óptima en el portafolio energético será diferente, y por ello, su planeación tiene que ser local.
Planear el cambio continuo de la mezcla energética no se reduce simplemente a determinar la proporción de cada una de las fuentes de energía en el portafolio, en los siguientes años. No se trata de determinar en qué cantidades vamos a comprar a otros países los satisfactores requeridos para producir la energía que habremos de demandar. Implica planear de qué manera vamos a producir nuestra propia energía renovable. Es decir, supone determinar los niveles de inversión requeridos, por ejemplo, para producir nacionalmente y a costos competitivos los módulos fotovoltaicos a instalar en nuestro país. Y esto a su vez, requiere determinar cómo se va a financiar ese desarrollo (inversión pública y/o privada), y qué número de personas capacitadas (ingenieros y técnicos) es necesario formar. O sea, tiene que ver con el establecimiento de una industria nacional que nos permita cierto grado de independencia tecnológica del extranjero, e incluso contar con una industria propia y que pueda ir más allá de solo satisfacer nuestras propias necesidades, y que pueda incluso llegar a ser exportadora de este tipo de productos, causando crecimiento económico. Un ejemplo de este tipo de planeación es la de un país como China, que hace sólo 10 a 15 años no contaba con una industria fotovoltaica, y ahora es el principal productor y exportador de módulos fotovoltaicos en el mundo. China es además el país con el mayor crecimiento de plantas generadoras de energía fotovoltaica. En el aspecto político-administrativo es necesario plantear las políticas públicas que ayudarían a fomentar el surgimiento de una industria nacional, y que nos permita resolver sinérgicamente nuestra demanda de energía eléctrica en el futuro a mediano y largo plazos (30 y 100 años, respectivamente).
Podemos resumir y concluir lo expuesto anteriormente: la energía solar fotovoltaica ya es económicamente competitiva con otras alternativas; en países con gran recurso solar (insolación) será inevitable que en el futuro una gran parte de la demanda energética (electricidad) sea satisfecha a través de esta fuente de energía renovable, y que además ayude a reducir la emisión de gases de efecto invernadero. Esto no se dará de manera automática si no se planea a mediano y largo plazos, tanto la mezcla energética como la forma en que se podrá alcanzar, sin ser dependientes de otros países, afirmando así nuestra seguridad energética. Habrá que responder ¿cómo se fomentará el surgimiento de una industria nacional?, ¿cómo se generará el personal capacitado?, ¿qué niveles de inversión se necesitan?, ¿qué políticas públicas deben plantearse? y ¿cuál es el desarrollo tecnológico y científico requeridos dentro de nuestras propias universidades para alcanzar las metas propuestas?. La participación multi-disciplinaria de diversos especialistas (economistas, legisladores, ingenieros, tecnólogos, planificadores financieros, etcétera) es esencial para establecer el plan para la transición energética. Sin políticas públicas, tanto para la transición energética como para el desarrollo industrial propio (en éste y otros sectores estratégicos), nuestro país estará condenado a seguir siendo dependiente, y con la pobreza endémica asociada a nuestro subdesarrollo industrial y tecnológico. ¿Será posible que haya un verdadero cambio en el futuro cercano?
BIBLIOGRAFÍA
Global Market Outlook for Solar Power 2023-2027, Solar Power Europe (2023).
Arturo Morales Acevedo (2016), “El Promisorio Futuro de la Energía Solar Fotovoltaica”, Avance y Perspectiva 2 (1), 50-53.
Arturo Morales Acevedo (2016-2017), “El mundo de los electrones en las Celdas Solares”, Avance y Perspectiva 2 (2), 50-53.
Una herramienta que dejó huella: 30 años de marcadores moleculares en plantas en el Cinvestav
El análisis genético depende de la capacidad de detectar diferencias (polimorfismos) entre individuos. Mendel por ejemplo, aprovechó las características visibles como tamaño, forma, color, entre otros rasgos (1). Más tarde algunos investigadores emplearon marcadores bioquímicos como proteínas u otros metabolitos como indicadores genéticos (2). Sin embargo, la mayoría de los caracteres estudiados se basó en mutaciones con efectos deletéreos, dificultando la posibilidad de dar seguimiento a varios caracteres que se presentan de manera simultánea.
Fundado en los avances en la metodología para manipular y visualizar fragmentos de ADN, Botstein (3) propuso un método apoyado en la detección de polimorfismos en la longitud de fragmentos de restricción, RFLP por sus siglas en inglés. Los RFLP se basan en diferencias en la capacidad de una enzima de restricción para reconocer y recortar una secuencia específica en el ADN. Este método tiene la ventaja que es neutral (no afecta físicamente al organismo del cual se extrae la muestra), y se sustenta directamente en el genotipo, evitando efectos ambientales o de la etapa de desarrollo del organismo, en la detección de los polimorfismos. A diferencia de un marcador fenotípico, como un marcador bioquímico, que tendría enormes variaciones dependiendo del tejido, etapa de desarrollo, y condiciones ambientales en los cuales se toma la muestra. Además, una misma metodología puede aplicarse a cualquier organismo (Figura 1). El desarrollo de los RFLP y un poco más tarde, los análisis de microsatélites o SSR (método basado en diferencias en el número de repeticiones de secuencias específicas de ADN, y empleado hasta la fecha para la identificación de individuos en casos criminales, de paternidad, etcétera), abrió la posibilidad de ampliar el análisis genético a organismos fuera de los modelos de estudio preferidos: maíz, ratón, Drosophila y empezar a correlacionar fenotipos con genotipos.
Figura 1. Dibujo que muestra el alcance de los marcadores moleculares para diferenciar entre individuos, en el caso de los humanos, y entre variedades cercanas, en el caso del maíz. Agradecemos al Dr. José Trinidad Ascencio Ibáñez (North Carolina State University) por su creatividad y talento en la preparación de esta figura.
Sin embargo, la metodología para detectar los polimorfismos RFLP o SSR es complicada, tardada y resulta difícil analizar varios marcadores simultáneamente. Con la innovación de la metodología de la reacción en cadena de la polimerasa (PCR), se buscó la manera de aprovechar la PCR para detectar polimorfismos (4). El primer método desarrollado basado en PCR, fue el de ADN polimórfico amplificado al azar (RAPD) que, aunque fue utilizado ampliamente, es difícil de replicar entre diferentes laboratorios (5).
En 1993 apareció una solicitud de patente por parte de la empresa Keygene establecida en Holanda para una nueva metodología combinando la detección de polimorfismos en sitios de restricción (igual que los RFLPs) con el uso de PCR (igual que los RAPD). Este procedimiento se conoce como polimorfismos en la longitud de fragmentos amplificados (AFLP), y además de las ventajas que ofrecen los RFLP, SSR y RAPD, la metodología de AFLP es robusta y facilita el análisis de cientos de marcadores a través del genoma de manera simultánea (6,7).
El método de AFLP se basa en la reducción del número de fragmentos de restricción que serán analizados. Si simplemente cortamos el ADN de un organismo con una enzima de restricción, y separamos los fragmentos por electroforesis en un gel, observaremos un barrido continuo de los cientos de fragmentos sin poder distinguir los fragmentos individuales, y aunque existen diferencias en la presencia de sitios de restricción entre distintos individuos, es imposible detectar estos polimorfismos. La metodología de AFLP permite la visualización de fragmentos individuales y su comparación entre diferentes individuos de manera sencilla.
Típicamente, el proceso experimental para los AFLPs se inicia con la digestión del ADN genómico del individuo usando dos enzimas de restricción tipo II distintas: una enzima que reconoce un sitio de seis pares de bases y otra enzima que detecta un sitio de cuatro pares de bases. En el siguiente paso se agregan adaptadores diseñados para unir específicamente a los sitios de restricción cortados por las enzimas y se lleva a cabo una reacción de PCR empleando oligonucleótidos con homología específica para los adaptadores (Figura 2). Dadas las condiciones de la reacción, los fragmentos con sitios de restricción para ambas enzimas son preferencialmente amplificados, mientras los fragmentos con sitios de restricción para una sola enzima no se amplifican de manera óptima. Así, se enriquece la muestra para los fragmentos con extremos portando sitios para diferentes enzimas. Sobre esta reacción enriquecida se lleva a cabo una segunda amplificación, con oligonucleótidos selectivos: los mismos que se utilizaron en la primera ronda de amplificación, pero ahora añadiendo 1 a 3 nucleótidos al azar extras hacia los extremos 3´ del fragmento de interés. De esta manera solamente los fragmentos que portan estos nucleótidos enseguida del sitio de restricción, serán amplificados, reduciendo de nuevo la complejidad del número de fragmentos y permitiendo su visualización después de su separación por electroforesis. Por cada nucleótido selectivo se reduce alrededor de 4 veces el número de fragmentos que serán detectados. La incorporación de nucleótidos marcados (por radiactividad o fluorescencia) en uno de los oligonucleótidos permite la visualización de los fragmentos de manera óptima (Figura 2).
Figura 2. Esquema que describe los pasos básicos del método de análisis de AFLP. Creado con BioRender.com
Es posible modificar la metodología de los AFLP para acomodar genomas pequeños como los de bacterias, reduciendo a 1 el número de nucleótidos selectivos o aumentándolos a 3 cuando el análisis involucra especies con genomas grandes como en el género Agave. Generalmente se amplifican entre 50 y 100 fragmentos, los cuales se separan por electroforesis en geles de acrilamida o en un secuenciador automático “tradicional” (secuenciación Sanger), donde se puede resolver fragmentos con diferencias de un solo nucleótido.
La Unidad Irapuato del Cinvestav fue el primer sitio en México donde se implementaron los métodos de análisis de marcadores moleculares tipo RFLP, RAPD y SSR en especies vegetales, específicamente en maíz y frijol, y el siguiente paso fue la incorporación de la metodología de los AFLP. Los colegas de Keygene generosamente permitieron acceso a la aplicación para la patente de los AFLP antes de su publicación formal (7), y con la infraestructura para la secuenciación de ADN ya existente en el Departamento de Ingeniería Genética de la Unidad Irapuato, fue relativamente sencillo instrumentar la metodología y atender el interés creciente para llevar a cabo trabajos de marcadores moleculares en plantas. La ubicuidad del método permitió aplicarlo a 60 especies vegetales, pero también en una amplia variedad de especies y géneros fuera del campo vegetal, incluyendo 11 especies de hongos, 3 de bacterias, 1 insecto y 1 especie de coral. La metodología de AFLP sentó las bases para un gran número de publicaciones y tesis de Licenciatura, Maestría y Doctorado del grupo de Genética Molecular de la Unidad Irapuato, que hasta la fecha ofrece servicios internos y externos de análisis de marcadores genético-moleculares incluyendo AFLP.
Las aplicaciones incluyen estudios de diversidad, por ejemplo en agave, piña y limón, entre otros; el desarrollo de mapas genéticos y asociación con caracteres de interés en maíz, frijol y varias especies de hongos y análisis de poblaciones de hongos patógenos. Algunos trabajos externos se han apoyado en la certificación de variedades y en resolver disputas de propiedad a nivel judicial con la determinación de diferencias o similitudes entre variedades comerciales de maíz y jitomate, producidos por distintas empresas de semillas. En casos de demandas específicamente con semillas de chile y melón, fue posible mostrar de manera inequívoca la venta de semillas apócrifas (Figura 3).
Figura 3. Dendrograma basado en AFLP utilizado para distinguir entre variedades legítimas y una variedad apócrifa de una especie vegetal. Las muestras C4 y C3 son controles de una especie vegetal de la misma familia que la especie de las variedades bajo análisis. Las muestras C1 y C2 son de la misma especie que las muestras bajo análisis. El esquema, árbol o dendrograma muestra la relación genética entre las diferentes muestras. Como se puede observar, las ramas A y B contienen las muestras apócrifas y legítimas respectivamente, indicando una clara diferencia a nivel genético. La longitud de las líneas verticales indica la distancia genética: mayor longitud = mayor distancia genética. Por ejemplo, la rama A que contiene muestras de la variedad apócrifa se separa de la rama B que contiene las variedades legítimas a una distancia aproximadamente de 0.08 mientras las variedades legítimas muestran un parentesco cercano con la separación de las ramas E y F en una distancia de 0.02 aproximadamente. Basado en estos datos podemos concluir que las muestras vendidas como variedades comerciales legítimas a un grupo de productores y que generaron rendimientos muy abajo del promedio, pertenecen en realidad a un genotipo (apócrifo) muy distinta a las variedades legítimas.
En colaboración con el Dr. José Ruíz Herrera (Q.E.P.D.), cuyo interés se enfocó en los cambios en patrones de metilación durante el dimorfismo en hongos, fue posible desarrollar una aplicación novedosa para los AFLP. Inicialmente los análisis de metilación estaban dirigidos hacia genes individuales, un trabajo laborioso que necesitaba conocimiento de las secuencias de los genes o regiones de interés del genoma. Adaptando la metodología de AFLP e incorporando enzimas de restricción sensibles a la presencia de nucleótidos metilados, fue posible extender la metodología de los AFLP para estudiar cambios en la metilación a través del genoma completo. Esta metodología se aplicó a estudiar cambios durante las diferentes etapas del dimorfismo de Mucor rouxii, Yarrowia lipolytica y Ustilago maydis (8). Es así como nació la metodología después conocida como MSAP-polimorfismos amplificados sensibles a la metilación (9). Aunque fue desarrollado originalmente en hongos, MSAP ha sido aprovechado ampliamente en especies vegetales, en Drosophila, y en mamíferos como humanos y ratón, entre otros (10).
Si bien AFLP y MSAP son utilizados ampliamente todavía (11, 12), la tendencia es reemplazar estas metodologías con nuevas estrategias para genotipificación basada en la Secuenciación de Nueva Generación (NGS). La reducción en costos para la secuenciación de genomas completos y el desarrollo de la metodología de “Genotyping by Sequencing o GBS”, basada en polimorfismos en nucleótidos individuales (single nucleotide polymorphisms), hace que la metodología de AFLP/MSAP ya no sea competitiva. En un sentido, el método de AFLP fue víctima de su propio éxito dado que los métodos de GBS emplean la misma estrategia de digestión con enzimas de restricción y amplificación de un número reducido de fragmentos, los cuales son secuenciados en lugar de ser separados por electroforesis (13, 14).
La historia de los AFLPs es un excelente ejemplo de cómo resolver un problema (reducir la cantidad de ADN para genotipificar) con una idea relativamente sencilla. Esta idea “sencilla” ha sido extendida para diferentes propósitos y ha tenido un impacto significativo con la publicación de por lo menos 300,000 artículos en revistas internacionales. Aunque el método original sin duda será obsoleto en los próximos años, el incremento acelerado en el uso de GBS asegura que la esencia del método de AFLP se mantendrá vigente al menos otros 30 años.
Referencias:
Mendel G. (1866). Versuche über Pflanzen-Hybriden Verhandlungen des naturforschenden Vereines in Brünn, 4, 3-47.
Brücher, H. (1989). Protein Plants. In: Useful Plants of Neotropical Origin. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-73313-0_4
Botstein, D., White, R.L., Skolnick, M., Davis, R.W. (1980). Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American journal of human genetics, 32(3), 314-331.
Mullis, K.B., Faloona, F.A. (1987). Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. In Methods in enzymology Vol. 155, 335-350. Academic Press.
Williams J.G.K., Kubelik A.R., Livak K.J., Rafalski J.A., Tingey S.V., (1990). DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18, 6531-6535.
Vos P., Hogers R., Bleeker M., Reijans M., Van de Lee T., Hornes M., Fijters A., Pot J., Peleman J., Kuiper M., Zabeau M. (1995). AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res 23:4407–4414.
Zabeau M., Vos P. (1993). Selective restriction fragment amplification: a general method for DNA fingerprinting. European Patent Application number: 92402629.7, Publication number 0534 858 A1.
Reyna-López, G.E. Simpson J., Ruiz-Herrera J. 1997 Differences in DNA methylation patterns are detectable during the dimorphic transition of fungi by amplification of restriction polymorphisms. Mol. Gen. Genet. 16, 847-852.
Zhao Y., Chen M., Storey K. B., Sun L., Yang H. (2015). DNA methylation levels analysis in four tissues of sea cucumber Apostichopus japonicus based on fluorescence-labeled methylation-sensitive amplified polymorphism (F-MSAP) during aestivation. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 181, 26-32.
Fulneček J., Kovařík A. (2014). How to interpret methylation sensitive amplified polymorphism (MSAP) profiles?. BMC genetics, 15(1), 1-9.
Panagiotopoulou H., Marzecki K., Gawor J., Kuhl H., Koper M., Weglenski P.,& Rzepkowska M. (2023). Extensive search of genetic sex markers in Siberian (Acipenser baerii) and Atlantic (A. oxyrinchus) sturgeons. Aquaculture, 573, 739517. 1-11.
Vogt G. (2023). Evolution, functions and dynamics of epigenetic mechanisms in animals. In Handbook of Epigenetics. Academic Press. 521-549.
Deschamps S., Llaca V., May G. D. (2012). Genotyping-by-Sequencing in Plants. Biology, 1(3), 460-483.
He J., Zhao X., Laroche A., Lu Z.-X., Liu H., Li, Z. (2014). Genotyping-by-sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Frontiers in Plant Science, 5, 484. 1-8.
Energía del desierto: descubriendo el nopal como fuente de biocombustible
Desde tiempos prehispánicos, el nopal se ha cultivado y consumido en México. En años recientes, su consumo ha aumentado debido a sus múltiples beneficios. Originario de las zonas secas de América, se ha extendido a todo el mundo. El nopal ha ganado popularidad debido a su valor nutricional y ventajas para la salud.
Este miembro de la familia de los cactus es una planta resistente y adaptable que cubre millones de hectáreas. Sus pencas miden entre 30 y 60 cm de largo, de 20 a 40 cm de ancho, y tienen un espesor de 1.9 a 2.8 cm. El cactus puede sobrevivir en ambientes secos y tierras infértiles, donde no prosperan otros cultivos (Figura 1).
La geografía y el clima de México brindan un ambiente propicio para su desarrollo. Con un rango de altitudes que va desde el nivel del mar hasta 2.600 metros y precipitaciones que llegan a 40 mm, el país ofrece las condiciones ideales. Los suelos arcillosos y ricos en sal complementan este entorno.
Un fenómeno de popularidad: del campo a la mesa
México lidera la producción y el consumo mundial de nopal. Su importancia histórica se remonta a los tiempos de las culturas prehispánicas , donde la economía y cultura se entrelazaron con esta planta. Los españoles, al descubrir su potencial, quedaron cautivados por ella. Incluso se utilizó para criar cochinillas, que generan un valioso tinte rojo, convirtiéndose en la tercera fuente de ingresos en la Nueva España. Hoy día, la demanda de cochinilla sigue siendo relevante.
Se piensa que el nopal acompañó a Colón en su viaje de regreso al Viejo Mundo. Las pencas se utilizaban en los barcos para prevenir el escorbuto en la tripulación, lo que ayudó a expandir su conocimiento y uso (Angulo y col., 2012).
Los nopales son sumamente nutritivos gracias a su composición química. Ésta puede variar según la especie, el medio ambiente, la cosecha y el procesamiento. Principalmente poseen agua (80-95%), carbohidratos (3-7%) y fibra (1-2%). Además, tienen bajos niveles de proteínas y grasas. En términos de minerales, son fuente de calcio y potasio, y también de cantidades pequeñas de magnesio, hierro y zinc. La fibra de los nopales está compuesta principalmente de celulosa.
Las raíces de estas plantas desempeñan un papel crucial en su supervivencia en lugares con escasez de agua. Para resistir la sequía, llevan a cabo algunas estrategias como limitar su área de exposición y reducir su tasa de evaporación.
Figura 1. El nopal en su hábitat natural.
Explorando los beneficios del nopal: una visión de un futuro saludable
Las plantas de las regiones áridas, como el nopal, evolucionaron para enfrentar la falta de agua. desarrollando el metabolismo tipo CAM (metabolismo ácido de las crasuláceas). En el metabolismo CAM, las plantas capturan dióxido de carbono durante la noche y lo almacenan en forma de ácido málico o ácido isocítrico en las vacuolas de las células y durante el día; la planta utiliza este almacenamiento de CO2 para llevar a cabo la fotosíntesis y producir carbohidratos. Los estomas de las plantas CAM permanecen abiertos durante la noche y cerrados durante la mayor parte del día, resultando de esta manera en una pérdida mínima de agua y fotorrespiración reducida. Este tipo de metabolismo es una adaptación eficiente para conservar agua en condiciones secas, ya que les permite realizar la fotosíntesis sin abrir completamente los estomas durante el día, reduciendo así la pérdida de agua por transpiración (Figura 2). Ejemplos de plantas que emplean el metabolismo CAM incluyen cactus, suculentas y algunas bromelias.
Figura 2. Esquema de metabolismo tipo CAM (metabolismo ácido de las crasuláceas) del nopal.
Además de sus pencas, se tienen los frutos del nopal – tunas – y sus semillas, valiosos en la industria alimentaria y farmacéutica. La semilla de tuna también puede ser una fuente para producir aceites o biodiésel. A pesar de su importancia, a menudo se le ha subestimado. Los extractos y las pencas también pueden contribuir a reducir los niveles de azúcar en la sangre en personas con diabetes.
El nopal es conocido con diferentes nombres alrededor del mundo. Por ejemplo, en Italia es “Fico d’India“, en francés “Raquette“, en portugués “Palma“, en turco “Sabra” y en Estados Unidos “prickly pear“. Estas variantes reflejan la fama del cactus y sus signficados diversos. Cada tipo de nopal presenta rasgos únicos según el lugar de crecimiento y las condiciones del suelo.
Figura 3. Estructura general de las betalaínas.
A medida que se aprende más sobre esta planta, se descubren más formas de aprovechar sus beneficios. Los nopales tienen flores de colores, como blanco, amarillo, rojo, naranja y rosa. A menudo estas flores tienden a ser desechadas una vez que se ha cosechado la fruta. A pesar de que se ha investigado menos sobre las flores en comparación con los frutos, se ha descubierto que contienen fenoles y pigmentos de betalaína (Figura 3), que son pigmentos naturales que se encuentran en muchas plantas, incluyendo el nopal. También tienen propiedades antioxidantes y antiinflamatorias. Estos compuestos solubles en agua son los causantes de los intensos colores. El nopal, con sus pencas, semillas y hojas, se convierte en un auténtico tesoro de la naturaleza.
Explorando nuevas fronteras: el nopal más allá de la mesa
El nopal no se limita a ser protagonista en la gastronomía mexicana. Su relevancia abarca más allá de la cocina, extendiéndose al campo de la energía. En la búsqueda de alternativas sostenibles, emerge con un gran potencial como fuente de biocombustibles. El elevado rendimiento y adaptabilidad lo sitúan como una opción energética viable. El nopal podría desempeñar un papel vital en la reducción de emisiones y la transición hacia fuentes más limpias.
Figura 4. Nopalera: un cultivo de nopales.
El uso de nuevas fuentes de energía en lugar de las tradicionales, requiere diferentes métodos y tecnologías. Entre las más destacadas se encuentran: solar, hidráulica, geotérmica, eólica, biomasa y bioenergía. La obtención de biocombustibles a partir de biomasa es uno de los métodos más empleados.
El biogás es un gas renovable obtenido a partir de la degradación anaerobia (sin oxígeno) de material orgánico. Este proceso de degradación biológico se realiza mediante la acción de bacterias que son sensibles o inhibidas por el oxígeno. El biogás está compuesto principalmente por dióxido de carbono y metano. La eficiencia de la digestión anaeróbica radica en la conversión de más del 90% de la energía disponible en metano, con un bajo consumo energético en el crecimiento bacteriano, en comparación con sistemas aeróbicos. Los microorganismos metanogénicos son fundamentales en este proceso, ya que actúan como enzimas respiratorias y son los encargados de la producción del metano.
Durante mucho tiempo la investigación energética en plantas CAM se centró en la producción de bioetanol. Sin embargo, el biogás ofrece ventajas significativas en comparación con el etanol. Su producción puede realizarse con tecnologías relativamente simples y en una gama moderada. A diferencia del etanol, que requiere plantas de procesamiento avanzadas y a gran escala, el biogás se separa fácilmente del agua, simplificando el proceso en comparación con las dificultades asociadas a la separación del etanol y el agua (Mason y col., 2015).
Casi el 90% de la biomasa de los nopales puede convertirse en biogás. El 10% restante se transforma en sedimento arrastrado por el agua que liberan los nopales. Este proceso genera residuos y agua como subproductos. El depósito rico en nutrientes puede reutilizarse como fertilizante.
El proceso inicia al introducir los nopales en biodigestores, donde las bacterias trabajan para producir biogás. Los nopales contienen mucha agua, alrededor del 88% en promedio. Además, los rendimientos de biogás son similares a los del maíz y otras biomasas. Este biogás es una mezcla de metano (75%) y dióxido de carbono (24%) que se utiliza como combustible y calefacción en los hogares.
En la búsqueda de energía limpia, se han realizado estudios sobre el rendimiento del nopal como fuente de biocombustibles. En particular, la producción de biogás a partir del nopal comenzó en Chile en el año 2000. El éxito técnico e industrial ha demostrado su viabilidad como alternativa sostenible para el futuro (Tabla 1).
Tabla 1. Comparación de biomasa de nopal con otras biomasas vegetales en términos de combustible y energía.
La búsqueda de una vida saludable: el nopal como aliado.
En México, desde 2016, una empresa llamada NopaliMex ha utilizado biogás de cactus para calefacción, combustible y electricidad; también produce biogás, fertilizantes y humus rico en nitrógeno. Tras purificar el biogás de dióxido de carbono, se convierte en biogás comprimido, empleado como combustible. Esta innovación no solo reduce costos, sino también disminuye la huella ambiental (Ciriminna y col., 2019).
Las capacidades caloríficas promedio varían según el nivel de dióxido de carbono. Al ser tratado con ácido sulfúrico para eliminar el dióxido de carbono y obtener biogás de alta pureza, llega al 96%. La cantidad de energía que se puede obtener del nopal depende de la producción de este último, su contenido de metano y rendimiento por hectárea.
Aunque las tierras semiáridas tienen un valor económico limitado, siempre existe algún área de oportunidad asociada a un cambio de uso. Con frecuencia se utilizan para pastoreo o ganadería de baja productividad. El empleo de estas tierras para biocombustibles puede aumentar la producción de alimentos a través de la agricultura bioenergética.
La Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO), estima que el 12,2% de la superficie terrestre del mundo es semiárida. Desde su papel histórico en la culturas prehispánicas, el nopal tiene una rica historia y un potencial prometedor que invita a explorar nuevos caminos hacia un futuro más verde y sostenible. Su uso en áreas secas puede preservar el agua y fomentar el desarrollo sustentable.
Conclusiones
El nopal no solo está arraigado en la historia de México, sino que también sobresale en el ámbito de la energía sostenible. Su capacidad para generar biogás y sus beneficios nutricionales han capturado la atención mundial. El nopal se presenta ahora como un elemento tanto en la cocina como en la energía. Adaptado para prosperar en climas secos, el nopal promete un futuro alentador. El nopal es un aliado a corto, mediano y largo plazo, cuando la sustentabilidad es pieza central.
Referencias
Mason P. M., Glover K., Smith J. A. C., Willis K. J., Woods J., Thompson I. P. “The potential of CAM crops as a globally significant bioenergy resource: moving from ‘fuel or food’ to ‘fuel and more food.” Energy Environ. Sci., 8, 2015, 2320-2329. DOI: 10.1039/c5ee00242g.
Ciriminna, R., Chavarría-Hernández, N., Rodríguez-Hernández, A.I. and Pagliaro, M. (2019), Toward unfolding the bioeconomy of nopal (Opuntia spp.). Biofuels, Bioprod. Bioref., 13: 1417-1427. https://doi.org/10.1002/bbb.2018
Angulo-Bejarano P. I., Paredes-López O., Nopal: A Perspective View on Its Nutraceutical Potential. Hispanic Foods: Chemistry and Bioactive Compounds. American Chemical Society Simposium Series. January 1, 2012, 113-159. DOI:10.1021/bk-2012-1109.ch009.