

Resumen
La evolución ha generado una diversidad extraordinaria de hongos. Estos pueden desarrollarse como organismos unicelulares o multicelulares, es decir, pueden crecer como una sola célula o formar estructuras complejas llamadas cúmulos, micelio, o cuerpos fructíferos, mediante la adhesión o agregación clonal de sus células. Estas estructuras les permiten transportar nutrientes, comunicarse entre células, e incluso diferenciarse para realizar funciones específicas. Además, estas características convierten a los hongos en organismos modelo para el estudio de la multicelularidad, los mecanismos de diferenciación, y la organización celular en eucariotas. En este trabajo se describe la multicelularidad de los hongos, y se da un repaso a las estructuras multicelulares conocidas de estos organismos eucariontes.
Introducción
La inmensa diversidad de organismos que cubren nuestro planeta, desde bacterias microscópicas hasta mamíferos gigantes como las ballenas azules, es sorprendente. ¿Cómo fue que se formó tal diversidad de seres vivos? A lo largo de la evolución se ha desarrollado una transición escalonada desde organismos unicelulares, relativamente simples, hacia organismos multicelulares mucho más complejos. A este proceso en el cual los organismos se agregan y cooperan para formar sistemas más complejos compuestos por muchas células se le llama multicelularidad. Según Krizsán y colaboradores (2019), esta transición ha ocurrido en distintos reinos biológicos, como plantas verdes, algas pardas, algas rojas, animales y hongos. La multicelularidad ha ocurrido en múltiples ocasiones a lo largo de la historia de la vida (Miller, 2010).
Al analizar la historia evolutiva de los hongos se percibe una transición celular interesante. Los primeros hongos que aparecieron y colonizaron ambientes acuáticos hace más de mil millones de años pudieron ser organismos unicelulares parecidos a los quitridiomicetos actuales (Nagy et al., 2018). Después de millones de años, estos hongos desarrollaron las células alargadas características llamadas hifas, que permitieron la colonización de ambientes terrestres. Esto propició la aparición de los primeros hongos que se asociaron a las plantas y formaron endomicorrizas, es decir, asociaciones de hongos y raíces en donde el hongo vive dentro de la célula vegetal. Además, entre las formas que fueron adquiriendo los hongos, están los hongos filamentosos saprófitos, que se alimentan de materia orgánica muerta. Más tarde aparecieron hongos con estructuras más complejas a las que llamamos “cuerpos fructíferos”, que son las formas de hongos más conocidas en la actualidad. La gran diversidad de aspectos y estructuras que se desarrollaron en los hongos provocó a su vez un alto nivel de diferenciación y organización celular. En la actualidad se pueden encontrar hongos unicelulares, multicelulares e incluso algunos capaces de alternar entre ambas fases con diferentes grados de complejidad.
¿Qué es la multicelularidad?
La multicelularidad puede definirse como una agregación de células que funcionan de manera organizada y especializada para realizar funciones complejas. La multicelularidad es el resultado de un fenómeno holístico que involucra la coordinación de procesos fisiológicos y morfológicos gobernados por diversos mecanismos moleculares. Un ejemplo fácilmente entendible de multicelularidad es la formación de tejidos, órganos y sistemas en animales y plantas. Las células que conforman un tejido poseen características particulares con una constitución fisiológica adaptada al tejido del cual forman parte. Otros ejemplos característicos de multicelularidad son los cuerpos fructíferos, mejor conocidos como setas u hongos, que es común encontrar en el campo durante la temporada de lluvias, cuando la humedad del ambiente y del suelo es alta (Figura 1).
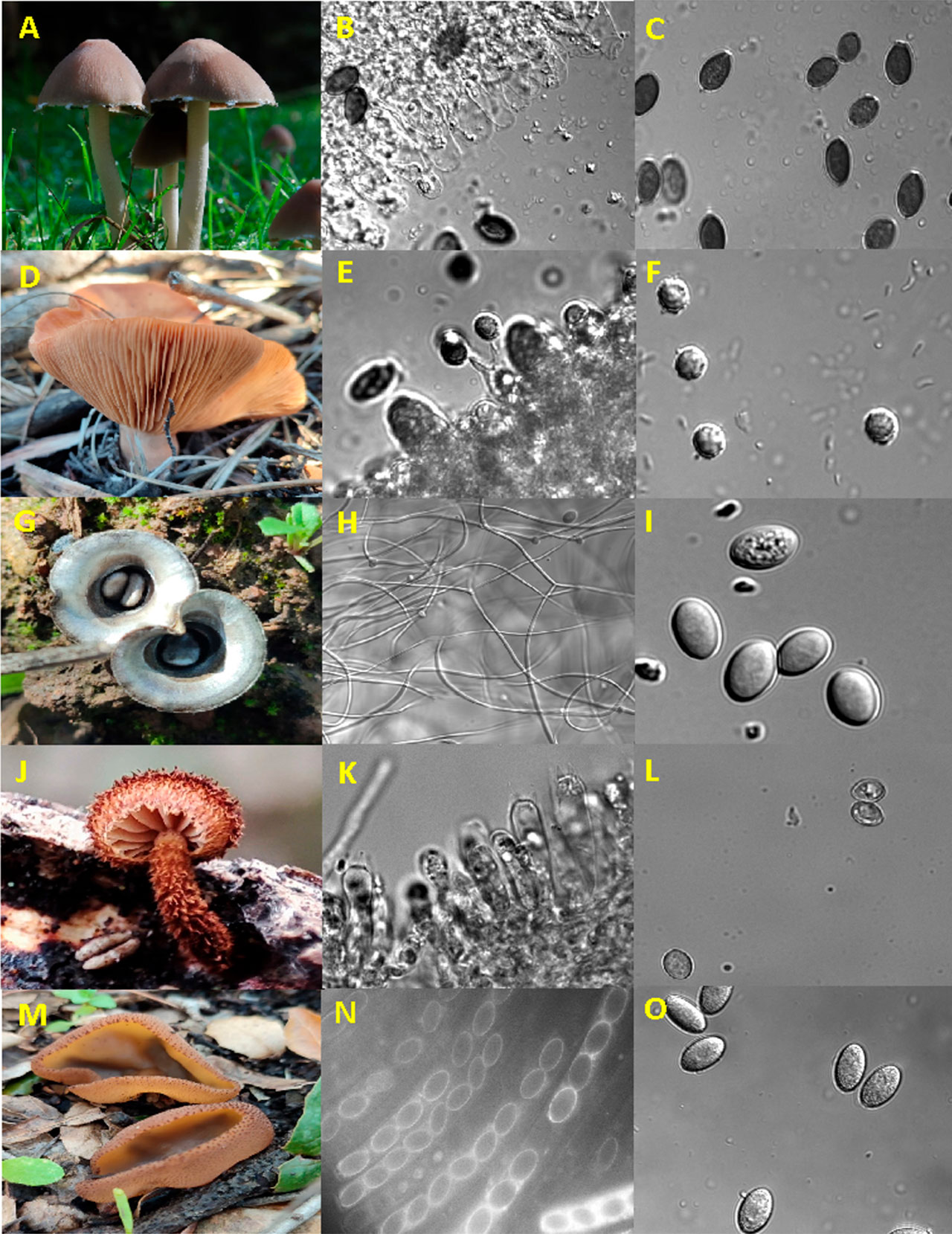
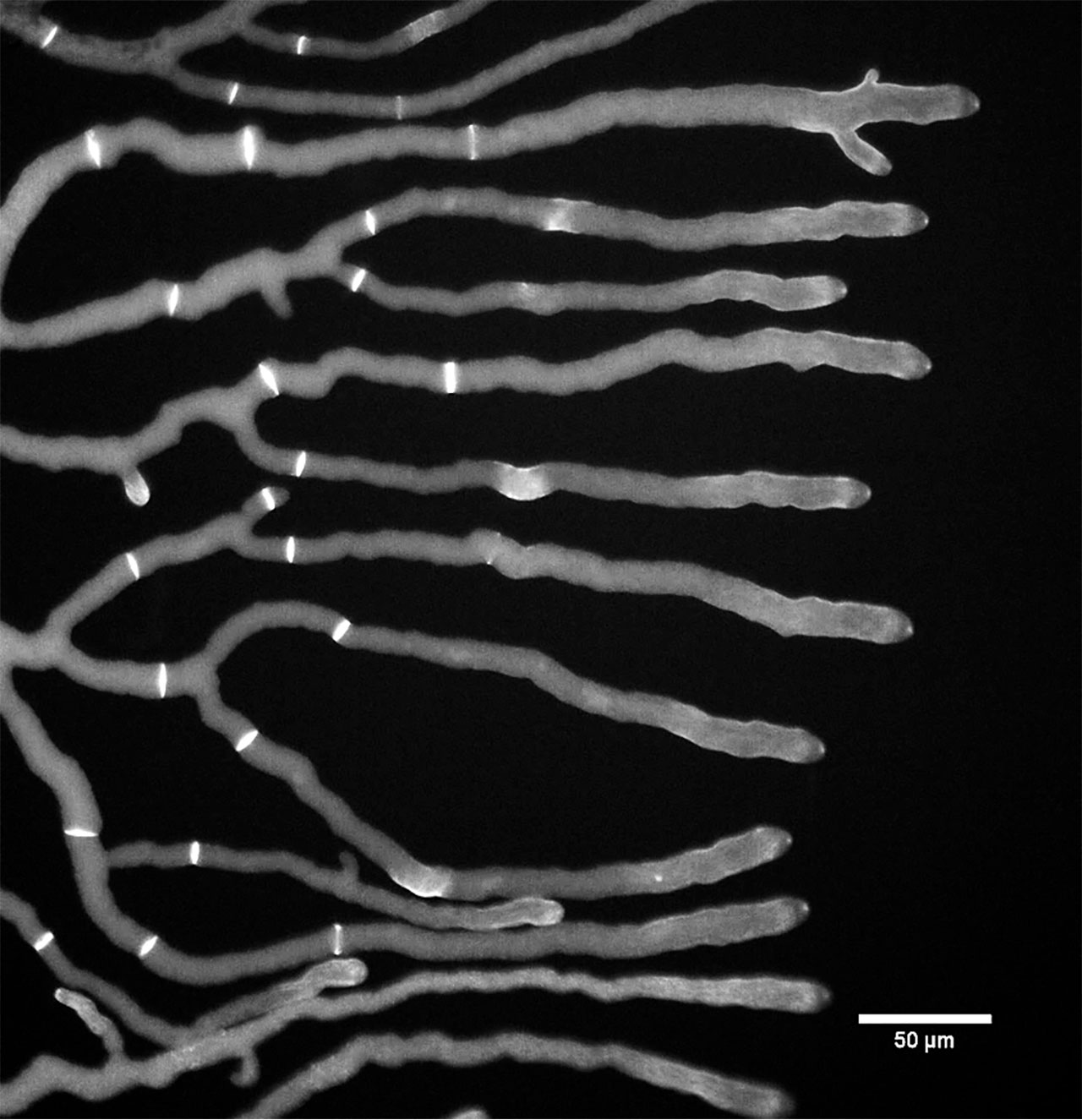
Los hongos pueden crecer de dos formas: como células individuales o filamentosas denominadas “levaduras” o “hifas”, respectivamente. En los llamados “hongos filamentosos”, la multicelularidad se presenta como micelio, que se compone por una red de hifas (Figura 2), por lo que son organismos sin rasgos definidos; la configuración del micelio es determinada por el sustrato en el cual vive. Las hifas son estructuras exclusivas del reino de los hongos. Se caracterizan porque crecen por la punta, son capaces de percibir su entorno y responder a cambios medioambientales (Martínez-Soto et al., 2017), de manera que pueden redistribuir agua y nutrientes a las zonas que lo requieran. Esta capacidad de crecer por la punta, de formar largos filamentos, de entrelazarse entre sí, ha significado que tengan una gran capacidad para colonizar ambientes, por ejemplo, sustratos muertos como madera, o capaces de invadir tejidos de organismos vivos. Por su parte, las levaduras han sufrido un proceso de “simplificación”, es decir, tienen una habilidad reducida para formar hifas, y pasan la mayor parte de su vida como formas unicelulares. Precisamente por esto, son menos conocidas como hongos multicelulares, aunque también desarrollan estructuras constituidas por múltiples células. En las levaduras, la multicelularidad se manifiesta como cúmulos celulares, llamados “clusters” en inglés (Figura 3), los cuales presentan una estructura y organización celular bien definida (Martínez-Soto et al., 2020a).
La multicelularidad de los hongos
Cúmulos celulares
Los hongos son sólo una de las ramas del árbol de la vida en la cual se desarrolló la multicelularidad. Aunque no sabemos del todo cómo empezó la vida, se sugiere que sus primeras manifestaciones fueron unicelulares. Es decir, las funciones vitales, estaban contenidas por completo en una sola membrana. Posteriormente, y después de los primeros hongos filamentosos, aparecieron las levaduras, las cuales muestran formas de organización multicelular tales como los cúmulos celulares (Figura 3). Éstos son grupos de células que se generan a partir de dos posibles mecanismos: mediante la agregación de células independientes para conformar una estructura multicelular quimérica, o por medio de la adhesión de las células madre e hija posterior a la mitosis para desarrollar una estructura multicelular que comparte el mismo genotipo (Martínez-Soto et al., 2020a).
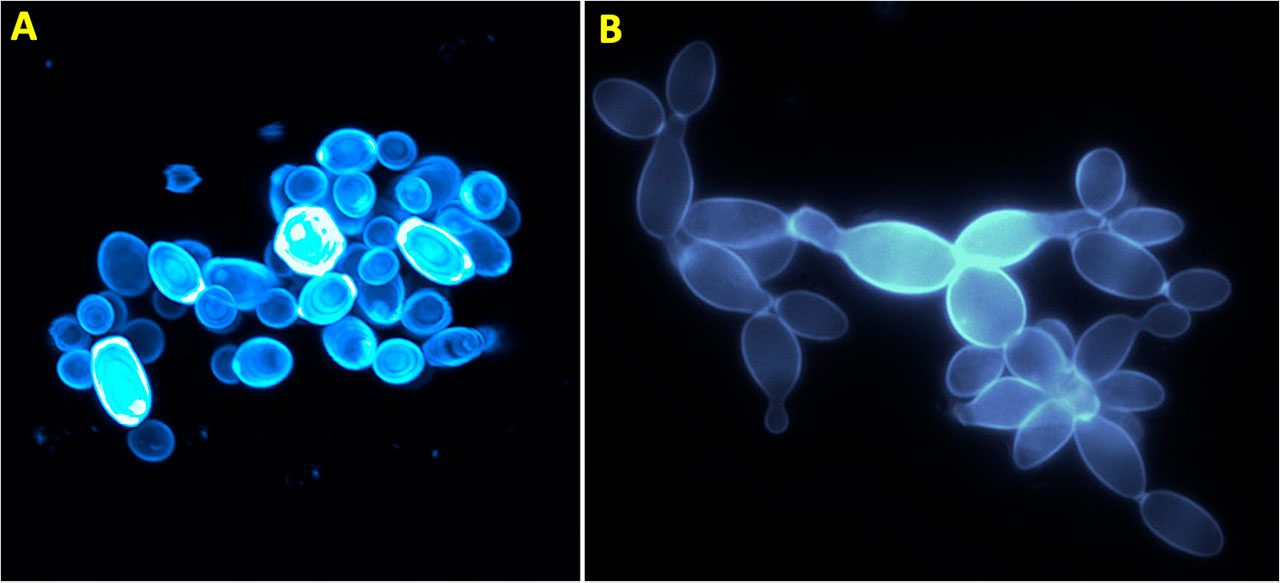
En algunos hongos, los cúmulos celulares generalmente se forman como respuesta directa ante diferentes estresores ambientales. Por ejemplo, se ha observado que el hongo Sporisorium reilianum, que parasita al maíz y al sorgo, forma cúmulos multicelulares con el fenotipo “copo de nieve” cuando se cultiva en medios con condiciones ácidas o en medios escasos de nutrientes (Martínez-Soto et al., 2020b) (Figura 3). Este mismo fenómeno se ha observado en la levadura Saccharomyces cerevisiae cultivada por largos periodos en biorreactores (Ratcliff et al., 2015). Diversos estudios revelan que cuando las estructuras “copo de nieve” alcanzan un tamaño considerablemente grande, la tensión entre sus células aumenta gradualmente, llegando a ser tan intensa que supera la resistencia de la conexión intracelular y provoca la eventual fragmentación del “cluster”. Esta ruptura, conocida como propagación celular, favorece la descendencia de los cuerpos multicelulares y es clave para la proliferación de estos microorganismos. Además, limita el posible conflicto genético al proporcionar nuevas mutaciones que promueven la adaptación y selección del organismo, como lo mencionan Ratcliff y coautores en su publicación del 2015.
De hifas a micelio
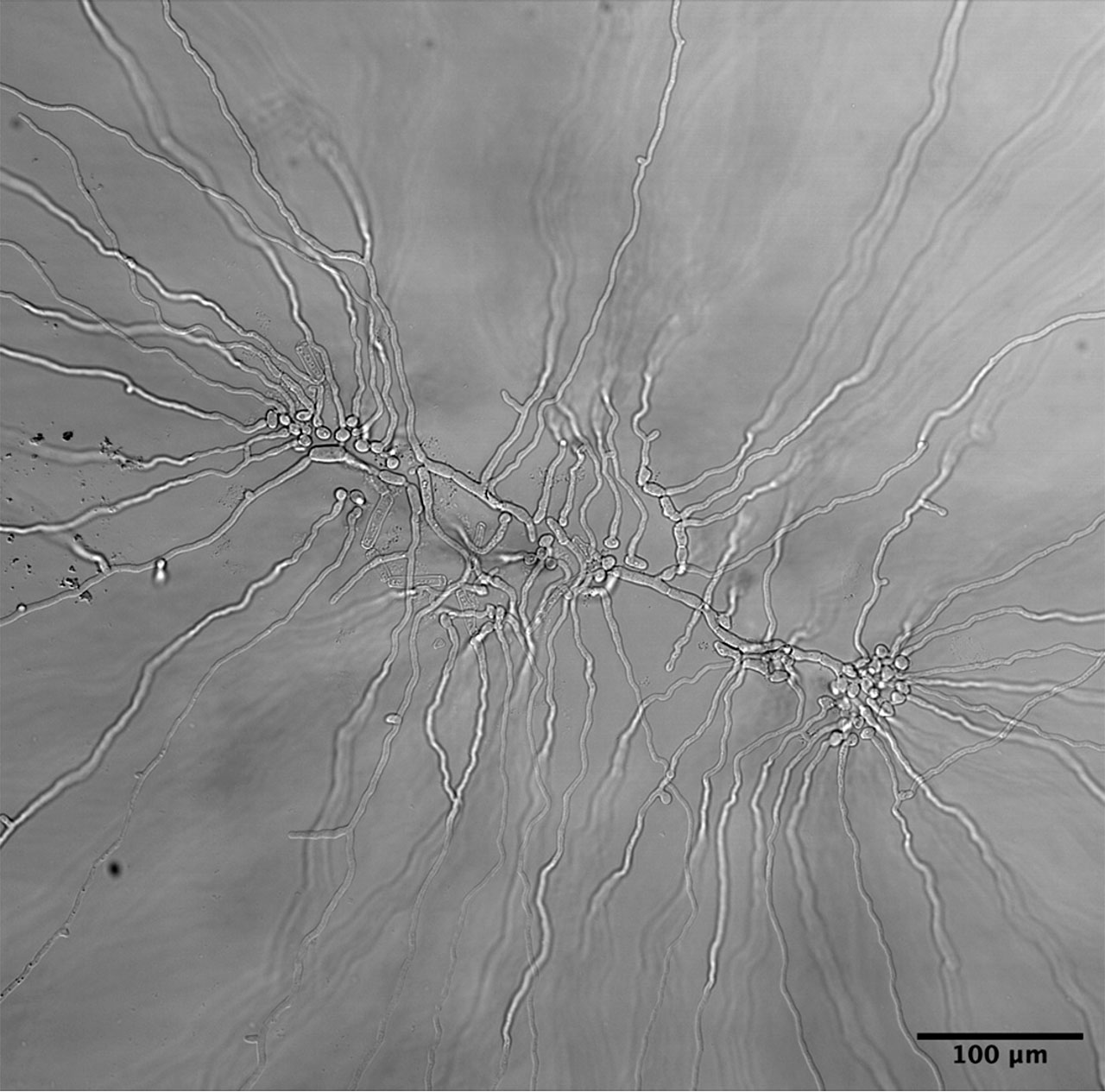
Las células de hongos como los mohos que crecen sobre alimentos olvidados en el fondo del refrigerador, o de frutos, o de cuerpos fructíferos, son denominadas “hifas”. De acuerdo con el diccionario etimológico de Anders (2023), la palabra “hifa” es un neologismo que proviene del latín moderno hypha (hyphae en plural), de la antigua voz griega hyphē (tejido o tela muy fina). La gran mayoría de los hongos conocidos están tejidos por esta fina tela celular, y pueden alcanzar formas muy diversas. La multicelularidad que caracteriza a los hongos proviene del crecimiento tipo fractal que exhiben las hifas (Figura 4).
El crecimiento hifal ocurre por medio de la extensión apical extremadamente polarizada, se logra por medio de la incorporación de membrana plasmática y los precursores de la pared celular en la punta de la hifa. A partir de esto, científicos como Nagy y colaboradores en 2020 se formulan la pregunta: ¿la hifa es la solución óptima para la multicelularidad de los hongos?, y consideran tres factores clave para el desarrollo de las hifas en los hongos: la evolución de la pared celular, el modo de nutrición a través de absorción, y la invasión de tierra.
Las interacciones mutualistas entre plantas y hongos fue un prerrequisito para la terrestrialización de las plantas, lo que a su vez requirió que los hongos fuesen capaces de obtener nutrientes en hábitats irregulares, en donde los nutrientes no se encuentran distribuidos de manera homogénea. La formación de cuerpos que se extendieron como tubos, con una pared rígida y el modo de nutrición por absorción, fue la solución al problema de la irregularidad de los nutrientes en este nuevo hábitat (Nagy et al., 2020). Siguiendo esta línea de pensamiento, se ha propuesto que el orden que pudo seguir el proceso evolutivo de los hongos multicelulares fue: el desarrollo del crecimiento apical, la compartimentación, la comunicación, diferenciación y adhesión celular para una formación posterior de los cuerpos fructíferos.
En la naturaleza los hongos se enfrentan con varios desafíos, como la competencia con otros organismos como bacterias, hongos y otros microorganismos. Por lo tanto, resulta favorable evitar estos retos y maximizar la eficiencia para adquirir nutrientes solubles. Los hongos se alimentan al absorber nutrientes producto de la digestión a través de enzimas que secretan al medio. Este es un proceso costoso, por lo que es muy importante que valga la pena producir las enzimas. Debido a los patrones de crecimiento apical y de ramificación del micelio, el cuerpo del hongo ocupa de la manera eficaz el área de sustrato en la que crece, por lo que se hace muy eficiente la absorción de estos nutrientes.
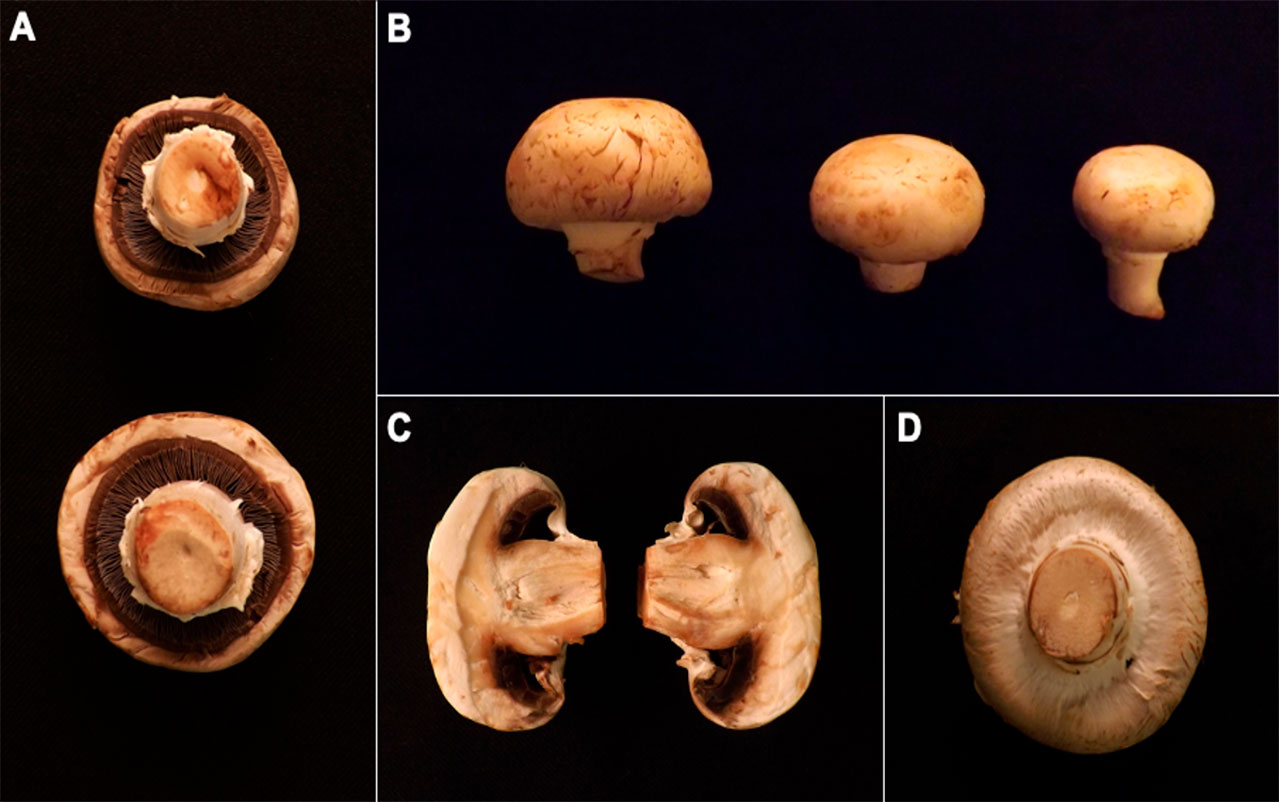
Es importante aclarar la diferencia entre un hongo y su cuerpo fructífero en sí. Como ejemplo citamos el caso más presente que tenemos de un hongo: el champiñón común. Esta estructura con sombrero y tallo se llama cuerpo fructífero, y es el sistema de reproducción del hongo. Tiene la función de producir y dispersar sus esporas sexuales. Los cuerpos fructíferos están compuestos por una multitud de células llamadas hifas especializadas en formar las esporas sexuales. Si se escarba un poco debajo del sustrato en el cual crece el cuerpo fructífero se encontrará una red con apariencia algodonosa llamada micelio. Podríamos decir que el micelio es el verdadero hongo, ya que está encargado de digerir y absorber los nutrientes del sustrato.
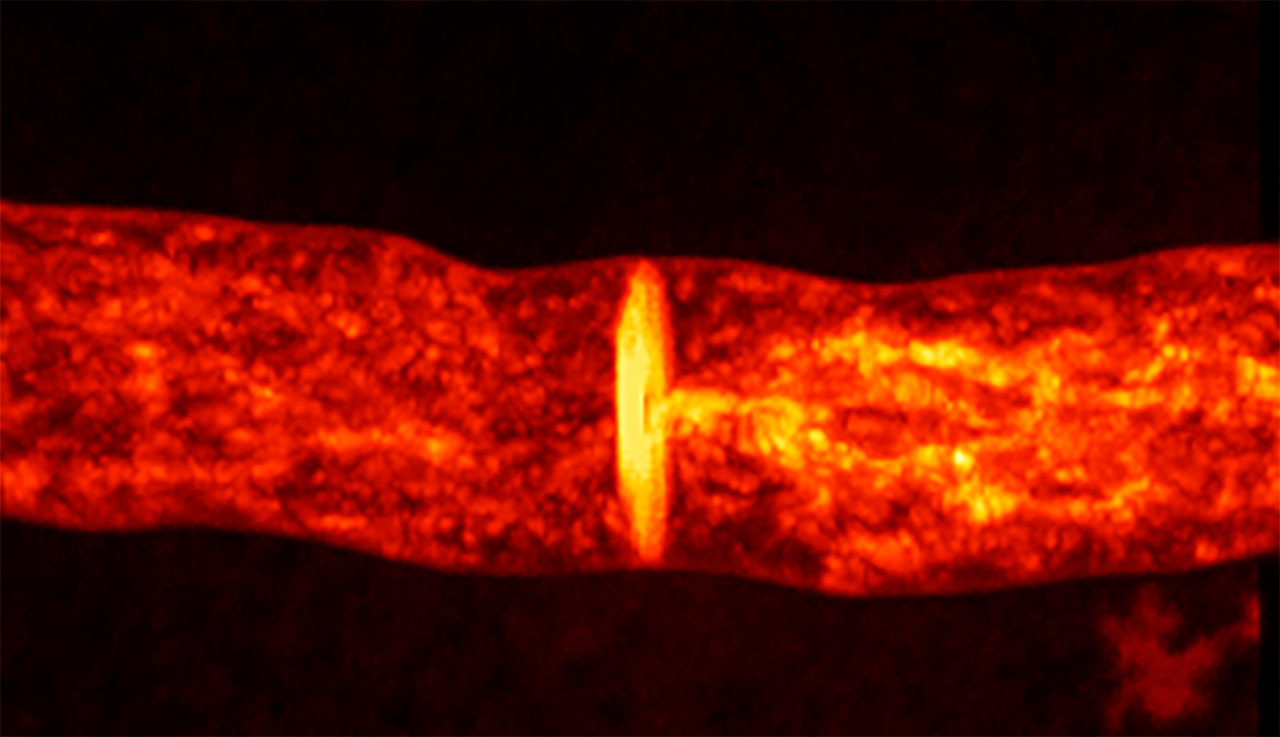
Los hongos escapan de la definición tradicional de un individuo. Si el micelio se llega a fragmentar, cada parte continuará creciendo de manera independiente y sin ningún límite más que el sustrato del que se alimenta. Los filamentos del micelio son llamados hifas y son las células del hongo. Una vez más, estamos frente a la dificultad de definir a una sola célula, porque estas hifas crecen, se expanden por la punta, ramifican y se fusionan (Figura 2). Aunque pueden formar paredes llamadas septos, que separan en compartimentos los filamentos de las hifas, al mismo tiempo se mantienen unidas permitiendo que el citoplasma y los organelos, incluyendo los núcleos, fluyan a través de los septos mediante aberturas llamadas poro septal (Figura 5). Distintos grupos de hongos pueden tener hifas sin septos, o septos con poros de complejidad variable que permiten mayor o menor flujo. En todos los casos, estos poros permiten la comunicación y el flujo entre compartimentos.
Las hifas de los hongos son células con una altísima presión osmótica. Para resistir esta gran fuerza, deben estar cubiertas por una pared celular fuerte. La maquinaria de crecimiento apical y de nutrición por absorción están acopladas: el mismo mecanismo que genera la expansión celular en la punta también se encarga de liberar las enzimas que digieren el sustrato extracelular para que pueda ser absorbido en forma de moléculas simples.
Las hifas, además de extenderse de forma eficiente, maximizan el uso del espacio tridimensional y de los sustratos que digieren y absorben. Podemos resumir que el cuerpo del hongo es el micelio, sin la capacidad de moverse (sésil), formado por un tejido de células con pared celular, que se alimenta por la absorción de moléculas simples. Bajo este entendido, resulta evidente la importancia de extenderse a lo largo de sustratos en los cuales los nutrientes no están distribuidos de manera uniforme, sino dispersos en forma de parches. De aquí la importancia de haber desarrollado un sistema de células largas, separadas, pero interconectadas para distribuir de manera eficiente los nutrientes a través de toda la red.
Los cuerpos fructíferos
Además del crecimiento ilimitado de las hifas que deriva en micelio de tamaño indefinido, los hongos que se reproducen sexualmente pueden desarrollar unidades multicelulares de tamaño definido, llamadas cuerpos fructíferos (un tipo de multicelularidad compleja). Aunque no está determinado el número preciso de tipos celulares diferentes que pueden formar los cuerpos fructíferos, se han definido 13 y 30 tipos distintos de células en los hongos Sordaria y Coprinopsis, respectivamente (Nagy et al., 2020).
A diferencia de otros linajes de vida que presentan multicelularidad, como animales, plantas, y algas, en los hongos la multicelularidad no inició por medio de adhesión de células. Es probable que ésta se haya desarrollado en los hongos para adherirse a materiales no-fúngicos como una forma de unirse a otros organismos o sustratos para infectarlos o alimentarse de ellos. A pesar de que la adhesión no fue crucial para establecer la multicelularidad en los talos formados por hifas, sí lo fue en algunas líneas fúngicas en las cuales se formaron otras estructuras multicelulares tridimensionales como los cuerpos fructíferos (Nagy et al., 2020).
Los cuerpos fructíferos de los hongos tienen una diversidad de forma muy acentuada. Como se mencionó, la estructura más conocida de un hongo probablemente es la de un champiñón (Figura 6). A esta forma se le llama “pileados-estipitados”, donde el píleo es a lo que se denomina “sombrero”, y el estípite es a lo que se llama tallo (Figura 6). Existen otras formas de hongos, como la forma de clava (o garrote), o la de corales, que se asemeja a los corales marinos. A estos hongos se les llama “clavarioides-coraloides”. También pueden tener píleos sin estípite, y quizás una de las formas más peculiares es la de los hongos gasteroides, que producen sus esporas internamente. Una de las ventajas fundamentales de la multicelularidad es que confiere a los organismos la posibilidad de dividir las labores entre los distintos tipos celulares. Así, en la formación de los cuerpos fructíferos, algunas células se especializan en producir esporas por medio de divisiones celulares continuas, mientras que otras lo hacen suministrando agua y nutrientes al resto del cuerpo fructífero.
En conclusión, se puede mencionar que los hongos son organismos clave para el entendimiento de la transición unicelular hacia la multicelular en eucariotas. Estos se han empleado para estudiar la importancia del desarrollo de estructuras especializadas y el papel que éstas juegan en incrementar su supervivencia. Considerando la diversidad de hongos existentes, así como sus innumerables formas y estructuras, aún falta realizar estudios que permitan elucidar los cambios genéticos y los mecanismos que, a través de la evolución, permitieron la generación de organismos multicelulares eucariontes mucho más complejos, como lo son los animales que conocemos hoy en día.
Referencias
Anders, V. et al., (2023). Etimología de televisión. Recuperado de https://etimologias.dechile.net el 2 de junio de 2023.
Krizsán, K., Almási, É., Merényi, Z., Sahu, N., Virágh, M., Kószó, T., … & Nagy, L. G. (2019). Transcriptomic atlas of mushroom development reveals conserved genes behind complex multicellularity in fungi. Proc. Natl. Acad. Sci. USA, 116, 7409-7418. doi: 10.1073/pnas.1817822116.
Martínez-Soto, D. y Ruiz-Herrera J. (2017). Functional analysis of the MAPK pathways in fungi. Rev Iberoam Micol. 34:192-202. doi: 10.1016/j.riam.2017.02.006.
Martínez-Soto, D., Ortiz-Castellanos, L., Robledo-Briones, M. y León-Ramírez, C. G. (2020a). Molecular mechanisms involved in the multicellular growth of Ustilaginomycetes. Microorganisms, 8, 1072. Doi: 10.3390/microorganisms8071072
Martínez-Soto, D., Velez-Haro, J.M., León-Ramírez, C.G. Galán-Vásquez E, Chávez-Munguía B, Ruiz-Herrera J. (2020b). Multicellular growth of the Basidiomycota phytopathogen fungus Sporisorium reilianum induced by acid conditions. Folia Microbiol 65, 511–521. https://doi.org/10.1007/s12223-019-00755-7
Miller, S. M. (2010) Volvox, chlamydomonas, and the evolution of multicellularity. Nature Education 3(9):65
Nagy LG, Kovács GM, y Krizsán K. (2018). Complex multicellularity in fungi: evolutionary convergence, single origin, or both?. Biol Rev Camb Philos Soc. 93:1778-1794. doi: 10.1111/brv.12418.
Nagy, L. G., Varga, T., Csernetics, Á., y Virágh, M. (2020). Fungi took a unique evolutionary route to multicellularity: Seven key challenges for fungal multicellular life. Fungal Biol. Rev. 34, 151-169. https://doi.org/10.1016/j.fbr.2020.07.002
Ratcliff, W. C., Fankhauser, J. D., Rogers, D. W., Greig, D., y Travisano, M. (2015). Origins of multicellular evolvability in snowflake yeast. Nat. Commun., 6, 6102. doi: 10.1038/ncomms7102.









